
Слои сетчатки не дифференцируются
Превращение исходного
зрительного пузырька
имеет и другое следствие. Если мы вспомним
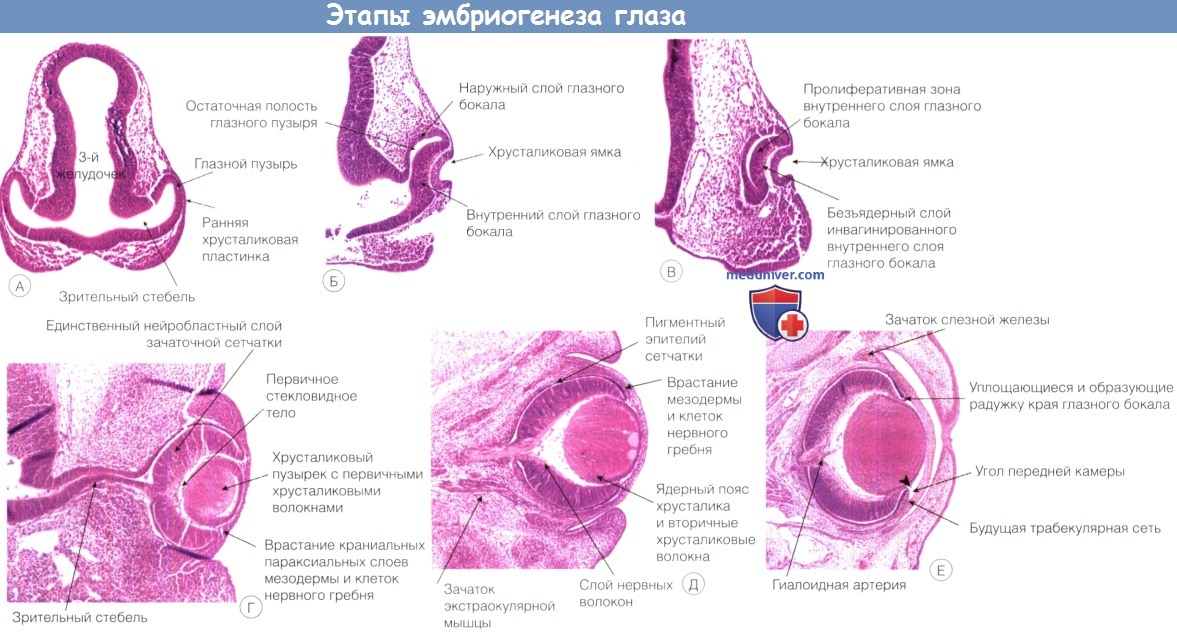
рис. 15.5
, мы увидим, что когда исходный зрительный желудочек исчезает, стенка
глазного бокала становится двухслойной. Наружный слой глазного бокала
формирует пигментный эпителий сетчатки, а внутренний — становится самой
сетчаткой. Хотя зрительный желудочек почти полностью закрывается в глазу
взрослого, соединение сетчатки с пигментным эпителием остается слабым
местом — отрыв сетчатки обычно происходит именно здесь. В норме сетчатка
удерживается на месте давлением стекловидного тела. Теперь слоям сетчатки
остается только дифференцироваться. Сетчатка — это, как мы подчеркивали
выше, «часть мозга, выставленная наружу». Из этого следует
предполагать, что ее дифференцировка сходна с таковой других частей мозга.
Цилиндрические эпителиальные клетки, выстилавшие исчезнувший зрительный
желудочек, преобразуются в герминативный слой. Первоначально число клеток в
этом слое невелико. Приблизительно на 4-й неделе развития наступает стадия
быстрой пролиферации, и число клеток здесь резко возрастает. После
завершения этой стадии клетки вступают в дифференцировку и перемещаются из
зрительного пузырька к
стекловидному телу
. Первыми дифференцируются ганглиозные клетки сетчатки. Они перемещаются
из герминативного слоя вдоль волокон, образуемых глиальными клетками
сетчатки — мюллеровыми клетками. По мере их перемещения из герминативного
слоя у них развиваются аксоны, идущие в составе зрительного нерва по ножке
к промежуточному мозгу. После этой первой волны дифференцировки из
герминативного слоя выделяется другой тип клеток — амакриновых. Возможно,
это изменение типа дифференцировки происходит вследствие секреции
ганглиозными клетками фактора, ингибирующего дальнейшую дифференцировку
герминальных клеток в ганглиозные. Амакриновые клетки, в свою очередь,
мигрируют из герминативного слоя к поверхности сетчатки, секретируя
вещества, ингибирующие формирование следующих амакриновых клеток. Вслед за
ними формируются биполярные, горизонтальные и промежуточные — и все они
покидают герминативный слой, двигаясь в сторону стекловидного тела и
дифференцируясь во все более сложном коктейле ингибиторов. Последними
дифференцируются фоторецепторные клетки — палочки и колбочки. Эти клетки не
мигрируют, а остаются по соседству с теперь уже виртуальным зрительным
пузырьком, опутанные отростками пигментных эпителиальных клеток. Теперь
клеточная структура сетчатки завершена, более подробно она будет
рассмотрена в
гл. СЕТЧАТКА ЧЕЛОВЕКА
. Стоит отметить, что не вся сетчатка формируется одновременно —
дифференцировка происходит в первую очередь на заднем полюсе глазного
бокала (дно), и только несколько недель спустя — на периферии. Вероятно
(хотя до сих пор остается объектом противоречий), что причиной больших
различий плотности клеток (особенно, ганглиозных) в разных частях сетчатки
является их избирательная гибель. Например, в фовеа ганглиозных клеток на
несколько сотен больше на единицу поверхности, чем на периферии сетчатки.
Наконец, стоит напомнить, что все эти сложные процессы оставляют
фоторецепторные клетки направленными внутрь — отвернутыми от падающего на
них света, да еще и покрытыми несколькими слоями нейронов. Это должно быть
головоломкой для тех, кто, считая эволюционные аргументы ниже своего
достоинства, во всем уповает на всемудрость Творца. Пока сетчатка и
хрусталик дифференцируются,
развиваются остальные части глаза
.
Смотрите также:
Источник
Эмбриогенез и дифференциация структур сетчатки, зрительного нерваИз двух слоев глазного бокала развиваются пигментный эпителий сетчатки и нейроретина. ПЭС необходим для роста глаза, он также участвует в процессе развития слоев нейроретины. Базальная пластинка ПЭС, мембрана Бруха, образует с пигментным эпителием и слоем хориокапилляров функциональную единицу. Она участвует в имеющем жизненно важное значение транспорте многочисленных биомолекул, кислорода, нутриентов и продуктов обмена между этими тканями. Дифференцировка нейроретины начинается в конце шестой недели. В любой зоне сетчатки соблюдается фиксированная последовательность развития клеток сетчатки. Ганглиозные клетки, горизонтальные клетки и колбочки появляются первыми, за ними следуют амакринные, биполярные клетки и палочки, последними появляются клетки Мюллера. На 10-11 неделе гестации выявляется участок наружного ядерного слоя центральной зоны сетчатки диаметром <500 мкм, примерно на 1500 мкм латеральнее диска зрительно нерва, содержащий только дифференцирующиеся колбочки — первый признак развивающейся центральной ямки. У приматов эти изменения являются отправной точкой самых значительных событий развития сетчатки, таких, как прекращение митозов, морфологическое созревание, апоптоз, экспрессия опсина и синаптогенез. Начиная с этой зоны те же процессы постепенно распространяются на всю сетчатку. Дифференцированный наружный ядерный слой — единственный слой, состоящий только из колбочек, сохраняющийся в центре фовеа у взрослого человека. Палочки и колбочки образуют правильную пространственную мозаику. Наибольшая пространственная плотность L- и М-колбочек отмечается в центре фовеа, тогда как S-колбочки отсутствуют в зоне 100 мкм от центра, а палочек нет в радиусе 300 мкм от центральной ямки взрослого человека. За несколько недель до рождения фоторецепторы всей сетчатки становятся морфологически более зрелыми. На 34 неделе палочки и колбочки вблизи исключительно колбочковой зоны фовеолярной области уже имеют наружные сегменты и формируют удлиненные аксоны, которые оканчиваются четко различимыми синаптическими ножками в сети отростков нейронов наружного плексиформного слоя. Хотя колбочки центральной ямки среди клеток сетчатки дифференцируются одними из первых, зрелые черты они приобретают последними. Для сравнения, колбочки периферии сетчатки достигают зрелости к перинатальному периоду. Генетическая абляция ПЭС или дизрупция генов специализации ПЭС у мышей приводит к микрофтальмии, трансдифференцировке ПЭС в сетчатку и появлению колобом. Васкуляризация сетчатки происходит путем пролиферации и миграции астроцитов и клеток эндотелия; с 14-15 недели гестации сосуды растут от диска зрительного нерва и примерно к моменту рождения достигают периферии сетчатки. Рост сосудов зависит от метаболической активности нейронов сетчатки, потребляющих кислород и вызывающих локальную гипоксию; таким образом стимулируется выработка астроцитами сетчатки эндотелиального сосудистого фактора роста (vascular endothelial growth factor — VEGF), который вызывает рост эндотелиоцитов и образование кровеносных сосудов сетчатки. После того, как сформированные сосуды начинают снабжать очередную область сетчатки оксигенированной кровью, выработка VEGF астроцитами снижается до базового уровня. Таким образом, формирование кровеносных сосудов определяется потребностью в кислороде. При нормальном развитии зона центральной ямки никогда не васкуляризируется, у человека фовеолярная аваскулярная зона полностью отграничивается примерно на 28 неделе гестации. Ингибирование роста сосудов сетчатки в центральной ямке вызвано появлением градиента (с центром в фовеа) антиангиогенных или антипролиферативных факторов, таких, как трансформирующий фактор роста — (3 или фактор роста фибробластов-2. В зоне центральной ямки сетчатка морфологически приспособлена к специфическим условиям кровоснабжения, возникающим из-за ингибирования развития сосудов сетчатки. Этими отличиями в строении фовеолярной области, возможно, объясняются предрасположенность макулы к развитию различных патологических изменений. Эмбриогенез зрительного нерваЗрительный нерв развивается из краев эмбриональной щели, в которой можно выделить два прилегающих друг к другу отдела: зрительную борозду, образующуюся из зрительного стебля, и ретинальную щель, возникающую на вентральной стороне глазного пузыря. Диск зрительного нерва образуется в зоне перехода зрительной борозды в ретинальную щель. В развивающемся диске зрительного нерва аксоны и гиалоидная артерия окружаются кольцом плотных нейроэпителиальных клеток, экспрессирующих содержащий парный бокс фактор транскрипции Рах2. Эти нейроэпителиальные клетки играют важную роль в развитии аксонов ганглиозных клеток (retinal ganglion cell-RGC) сетчатки. Первоначально аксоны ганглиозных клеток тянутся вдоль поверхности стекловидного тела по нейроретине от периферии к центру сетчатки. В самой сетчатке аксоны ганглиозных клеток растут в плотном контакте с подошвами глиальных клеток Мюллера и базальной пластинкой стекловидного тела, обе эти структуры вырабатывают молекулы клеточной адгезии и экстрацеллюлярного матрикса. Количество аксонов ганглиозных клеток, врастающих в зрительный нерв, достигает максимума в 3,7 миллионов примерно на 16-17 неделе гестации. Затем число аксонов уменьшается примерно до 1,1 млн к 29 неделе гестации. Эти числа совпадают с оценками количества аксонов в зрительном нерве взрослого человека в 1,1-1,3 миллионов. Избыточное количество аксонов участвует в формировании ретинотопической проекции коры посредством зависимых механизмов. Эмбриогенез других черепных нервовНа пятой и шестой неделях развития формируются двенадцать пар черепных нервов (cranial nerves — CN). Они подразделяются на три группы в зависимости от их эмбрионального происхождения. Соматические эфферентные черепные нервы: глазодвигательный (CNIII — большая часть нерва), блоковый (CNIV), отводящий (CNVI) и подъязычный (CNXII). Нервы жаберных дуг: CN V, VII, IX и X. Чувствительные нервы: CNI, CNII и CNVIII. В головном мозге (в отличие от спинного), организация двигательных нейронов определяется строением нейромеров. Черепные двигательные нейроны располагаются в пределах отдельных ромбомеров (нейромеры ромбовидного мозга) или в смежных парах нейромеров. Ядро глазодвигательного нерва (III) локализуется в заднем отделе среднего мозга, ядро блокового нерва (IV) лежит в переднем ромбомере 1 (r1), ядро тройничного нерва (V) — в ромбомерах 2 и 3, ядро отводящего нерва (VI) — в ромбомерах 4 и 5, ядро лицевого нерва (VII) — в ромбомерах 5 и 6, ядра языкоглоточного нерва (IX) — в ромбомерах 6 и 7. Средний мозг и каждый сегмент заднего мозга имеют свой молекулярный «адрес», проявляющийся экспрессией специфической комбинации факторов транскрипции. Нормальное развитие и функционирование эфферентных черепных нервов имеет решающее значение для нормального развития экстраокулярных мышц. Стромальный клеточный фактор-1 (stromal cell-derived factor-l-SDF-1) и фактор роста гепатоцитов (hepatocyte growth factor-HGF) играют важную роль в развитии аксонов глазодвигательного и блокового нервов. SDF-1 и HGF экспрессируются мезенхимой вокруг точек выхода нервов. SDF-1 и HGF также экспрессируются вокруг экстраокулярных мышц и ускоряют периферический рост аксонов глазодвигательного и блокового нервов, определяя их участие в формировании нервно-мышечных проекций.
— Также рекомендуем «Эмбриогенез и дифференциация глазодвигательных мышц, слезных органов» Оглавление темы «Эмбриология глаза»:
|
Источник
Пигментный ретинит (ПР), или тапеторетинальная абиотрофия (генерализованная наследственная дегенерация сетчатки, периферическая дистрофия сетчатки) является тяжелым наследственным прогрессирующим заболеванием органа зрения с первичным генетически детерминированным поражением фоторецепторного слоя и пигментного эпителия сетчатки (ПЭС).
Возникновение и развитие заболевания возможно в любом возрасте. Особенностью процесса является незаметное начало, сопровождающееся постепенным снижением зрительных функций. Наследуется по аутосомнотдоминантному, аутосомно-рецессивному и Х-сцепленному типу наследования, а также отмечена связь ПР с митохондриальными патологическими процессами [Boughman J.A. et al., 1980; Bunker C.H. et al., 1984; Bird A.C., 1988].
Клиническое течение пигментного ретинита характеризуется снижением темновой адаптации, нарастающим сужением поля зрения и угасанием показателей электрической активности сетчатки. На глазном дне обнаруживается типичное скопление пигмента в форме «костных телец» или, реже, мелких глыбок, зерен, локализованных преимущественно вблизи экватора. Часто встречается та или иная патология желтого пятна, возникает нарастающее сужение сосудов сетчатки, атрофические изменения цилиарного тела и радужной оболочки.
Гистоморфологически при пигментной дистрофии обнаруживается деструкция палочек и колбочек с вакуолизацией их наружных члеников. Наблюдается набухание, десквамация и смещение во внутренние слои сетчатки клеток пигментного эпителия, исчезновение ядерных и плексиформных слоев. Имеют место разрастание глии, фиброз, гиалиноз ретинальных и хориоретинальных сосудов, атрофия сосудистой. Область с костными тельцами не имеет фоторецепторов, содержит частично сохраненные клетки пигментного эпителия, интактную наружную пограничную мембрану. Наблюдаются друзоподобные дегенеративные изменения на уровне мембраны Бруха [Weleber R., 1998].
Морфологические изменения начинаются с миграции ядра от наружного нуклеарного слоя к слою палочек и колбочек и наружному плексиформному слою, что приводит к сморщиванию фоторецепторов в результате дегенерации целого комплекса клеток [Cartner S., Henkind P., 1982]. В ранних и промежуточных стадиях пигментного ретинита с аутосомно-доминантной формой наследования колбочки сохраняются дольше, чем палочки. Изначально нормальные фоторецепторы изменяются, их наружные сегменты расширяются и укорачиваются.
Патологический процесс начинается в экваториальной зоне и распространяется к центру и на периферию, при этом он более выражен в нижних квадрантах. В развитых стадиях заболевания наружные слои сетчатки полностью дегенерированы, исчезают фоторецепторы, волокна в наружном плексиформном слое и частично ядра внутреннего нуклеарного слоя и ганглиозных клеток. Клетки ПЭС изменены в областях, где фоторецепторы отсутствуют. Они могут мигрировать в нейрональные слои сетчатки или оседать в наружной пограничной мембране. В зонах с сохраненными фоторецепторами клетки ПЭС остаются нормальными.
Оценить состояние хориоретинального комплекса и тяжесть развития заболевания возможно с помощью ОСТ. На томограммах оценивается толщина слоя фоторецепторов, нервных волокон и нейроглии сетчатки, прозрачность слоев сетчатки относительно стандартной цветовой шкалы прибора, состояние ПЭС и слоя хориокапилляров. Эти изменения хориоретинального комплекса и были положены в основу предложенной классификации [Жукова С.И. и соавт, 2004].
В латентной стадии ПР, выявленной у 31 % обследованных ближайших родственников больных ПР при отсутствии клинических проявлений и офтальмоскопических признаков заболевания, обнаруживаются характерные изменения хориоретинального комплекса: уменьшение толщины слоя фоторецепторов, которое является следствием уменьшения длины наружных сегментов, либо исчезновением клеток и подтверждает предположение о первичном поражении фоторецепторов; снижение прозрачности данного слоя; увеличение толщины слоя ПЭС, обусловленное усиленным обновлением наружных сегментов и повышенным метаболизмом пигментного эпителия. Выявленные изменения подтверждают наше предположение о вовлечении в патологический процесс центральных отделов сетчатки уже на ранних стадиях заболевания (рис. 1).
, слой пигментного эпителия (2) расширен.")
Рис. 1. ОСТ макулы больного пигментным ретинитом в латентную стадию заболевания. Снижение прозрачности и уменьшение толщины слоя нейроэпителия (1), слой пигментного эпителия (2) расширен.
В стадии первых клинических признаков и прогрессирования дистрофических процессов в хориоретинальном комплексе выявляется дальнейшее снижение прозрачности слоя фоторецепторов (рис. 2) и уменьшение его толщины. Наблюдается неравномерность толщины слоя ПЭС с появлением в нем единичных дефектов, уменьшение толщины слоя хориокапилляров. Появление высокорефлектирующего ярко-красного окрашивания сетчатки обусловлено образованием пролиферативной глиальной ткани, которая формируется в результате миграции ПЭС, при этом усиленно продуцируются фибробласты и избыточно образуется рубцовая ткань, формируя на поверхности сетчатки эпиретинальную мембрану.
, появились дефекты ПЭС (2), разрушение слоя хориокапилляров (3).")
Рис. 2. Макула пациента больного ПР в стадию первых клинических признаков.
А. Биомикроскопия глазного дна. Типичные для ПР изменения отсутствуют.
Б. ОСТ изображение: отмечается разрушение слоя нейроэпителия (1), появились дефекты ПЭС (2), разрушение слоя хориокапилляров (3).
В стадии манифестации клинических проявлений ПР и генерализации дистрофических изменений в хориоретинальном комплексе отмечается отсутствие признаков дифференциации слоев сетчатки, формирование фиброзного слоя на внутренней поверхности сетчатки (рис. 3). В местах скопления пигмента фото-рецепторы отсутствуют, сетчатка резко истончена (рис. 4). У трети пациентов в данную стадию выявляется кистозный отек в центральных отделах сетчатки (рис. 5, 6).

Рис. 3. Макула больного ПР в стадию манифестации клинических признаков.
А. Биомикроскопия глазного дна. В экваториальной зоне видны скопления пигмента в виде «костных телец».
Б. ОСТ-изображение: слои сетчатки почти не дифференцируются. Красное окрашивание сетчатки указывает на ее глиальное перерождение.

Рис. 4. ОСТ средней периферии сетчатки больного ПР в стадию манифестации клинических проявлений. Скан сделан на участке скопления костных телец. Видно, что в месте пигментации сетчатка резко истончена, нейроэпителий отсутствует.

Рис. 5. ОСТ макулы пациента с ПР в стадии манифестации клинических проявлений. Слой фоторецепторов плохо дифференцируется. Крупные интраретинальные кисты в макуле. В отличие от других видов кистовидного отека обращает на себя внимание уплотнение внутренних слоев сетчатки.

Рис. 6. ОСТ макулы пациента с ПР в стадии манифестации клинических проявлений. Обширный интраретинальный крупнокистозный отек. Прослеживаются фрагменты разрушенных Мюллеровских клеток. Разрушение и перерождение всех слоев центральных отделов сетчатки.
Дегенеративные изменения достигают своего пика в стадии угнетения функций зрительной системы и тотальной деструкции хориоретинального комплекса (рис. 7).

Рис. 7. Макула пациента с ПР в стадии угнетения функций зрительной системы.
А. Биомикроскопия глазного дна. Нарушение ПЭС по всей площади глазного дна с формированием участков «голой» хориоидеи. ДЗН восковидно-бледный.
Б. ОСТ макулы. Слои сетчатки не дифференцируются. Наблюдается резкое истончение фоторецепторов и слоя хориокапилляров. Отмечается разрастание пролиферативной глиальной ткани. Выявляются обширные дефекты пигментного эпителия.
Изменения диска зрительного нерва при пигментном ретините характеризуется как нисходящая атрофия зрительного нерва, возникающая при поражении периферического нейрона зрительного пути, и медленно доходящая до диска.
В ранних стадиях заболевания изменений со стороны ДЗН не выявлено. В развитых стадиях отмечается типичная «восковидная» атрофия. Исчезновение слоя нервных волокон при ПР рассматривают, как результат транссинаптической дегенерации или первичного поражения фоторецепторов наряду с другими нейронами сетчатки (рис. 8).

Рис. 8. Диск зрительного нерва больного с ПР.
А. Биомикроскопия глазного дна. ДЗН уменьшен в размере, форма диска атипичная — он овальный, неправильной формы, имеет двухконтурный край; бесструктурен, границы диска несколько смазаны. Наблюдается своеобразное монотонно-желтоватое окрашивание диска, сочетающееся с его выстоянием. Количество нисходящих с диска сосудов уменьшено, артерии и вены сужены, выход сосудистой воронки с диска смещен к периферии, количество сосудов на сетчатке и ветвление их уменьшены, просвет заметно сужен.
Б. ОСТ ДЗН. Характерное выстояние диска зрительного нерва в стекловидное тело; атипичное расположение экскавации; разрастание соединительной ткани; снижение прозрачности структур.
Нередко при ПР обнаруживаются друзы ДЗН. Гистологически они представляют собой отложения гиалина и локализуются впереди решетчатой пластинки (рис. 9). Этиология друз неясна. Высказываются предположения об их наследственной природе.

Рис. 9. Друзы диска зрительного нерва у больного с ПР.
А. Биомикроскопия глазного дна. Небольшие округлые множественные очажки размером 1—2 диаметра вены сероватого цвета в краевой зоне ДЗН.
Б. ОСТ того же пациента. Оптически плотные округлые образования по краю ДЗН.
Таким образом, представленные материалы подтверждают, что оптическая когерентная томография сетчатки является объективным методом диагностики, как ранней стадии развития ПР, так и дополнительным критерием для классификации клинически выраженных стадий заболевания, позволяет осуществлять мониторинг патологического процесса и может быть использована в диагностике ПР, включая беспигментную форму, в том числе и у детей, когда из-за маленького возраста ребенка и его неадекватного поведения, невозможно проведение функциональных методов исследования.
Оптическая когерентная томография в офтальмологии
под ред. А.Г. Щуко, В.В. Малышева
Опубликовал Константин Моканов
Источник

