
Рецепторный потенциал в сетчатке
Оглавление темы «Рецепторный потенциал палочек и колбочек. Рецептивные поля клеток сетчатки. Проводящие пути и центры зрительной системы. Зрительное восприятие.»:
1. Рецепторный потенциал палочек и колбочек. Ток ионов через мембрану фоторецептора в темноте и на свету.
2. Адаптация фоторецепторов к изменениям освещенности. Световая адаптация. Десенситизация. Темновая адаптация.
3. Рецептивные поля клеток сетчатки. Прямой путь передачи сигналов от фоторецепторов к ганглиозной клетке. Непрямой путь передачи сигналов.
4. Рецептивные поля с оn-центрами и off-центрами. On-нейроны. Off-нейроны. Ганглиозная клетка on-типа. Ганглиозная клетка off-типа.
5. Рецептивные поля цветового восприятия. Восприятие цвета. Первичные цвета. Монохромазия. Дихромазия. Трихромазия.
6. М- и Р-типы ганглиозных клеток сетчатки. Магноцеллюлярные (М-клетки) клетки. Парвоцеллюлярные (Р-клетки) ганглиозные клетки сетчатки.
7. Проводящие пути и центры зрительной системы. Зрительный нерв. Зрительные тракты. Глазодвигательный рефлекс.
8. Латеральное коленчатое тело. Функциональная организация латерального коленчатого тела. Рецептивные поля латерального коленчатого тела.
9. Переработка зрительной сенсорной информации в коре. Проекционная зрительная кора. Световая грань. Комплексные нейроны. Двойные противоцветные клетки.
10. Зрительное восприятие. Магноцеллюлярный путь. Парвоцеллюлярный путь. Восприятие формы, цвета.
Рецепторный потенциал палочек и колбочек. Ток ионов через мембрану фоторецептора в темноте и на свету.
Специфической особенностью фоторецепторов является темновой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуанозинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фосфодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается.
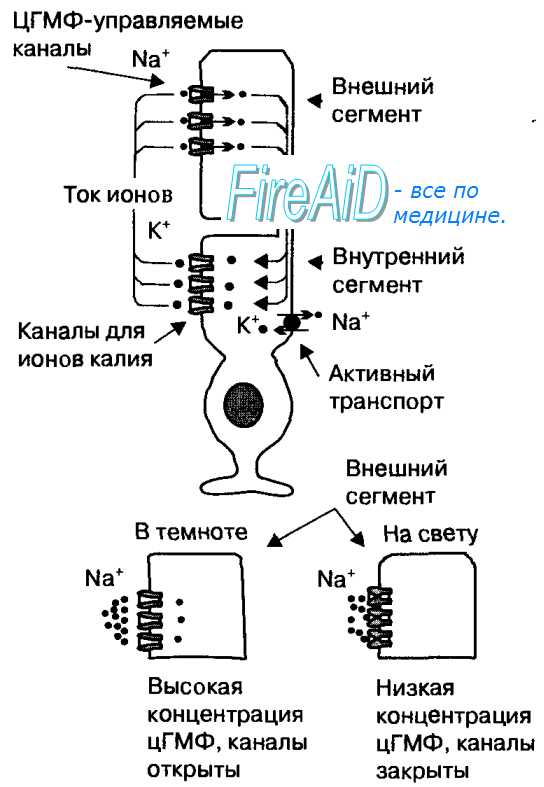
Рис. 17.8. Ток ионов через мембрану фоторецептора в темноте и на свету.
Ионы натрия поступают в фоторецепторы через катионные каналы внешнего сегмента, которые открываются при высокой концентрации цГМФ в темноте (темновой ток). Поступление в клетку ионов натрия вызывает деполяризацию мембраны фоторецептора до —40 мВ. На свету концентрация цГМФ уменьшается, что приводит к закрытию катионных каналов. Непрекращающийся выход ионов калия ведет к гиперполяризации мембраны, которая представляет собой рецепторный потенциал.
Вследствие непрерывного выхода ионов калия из клеток, мембрана фоторецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
Фоторецепторы образуют синапсы с биполярными клетками двух типов, различающихся по способу управления хемозависи-мыми натриевыми каналами в синапсах. Действие глутамата приводит к открытию каналов для ионов натрия и деполяризации мембраны одних биполярных клеток и к закрытию натриевых каналов и гиперполяризации биполярных клеток другого типа. Наличие двух типов биполярных клеток необходимо для формирования антагонизма между центром и периферией рецептивных полей ганглиозных клеток.
— Также рекомендуем «Адаптация фоторецепторов к изменениям освещенности. Световая адаптация. Десенситизация. Темновая адаптация.»
Источник
Мы познаем окружающий нас мир при помощи органов чувств. Эта уникальная способность связана с наличием специальных клеток – рецепторов, чувствительных к тем или иным видам воздействий, таким как свет, звук, запах, прикосновение и др. Рецептор– это высокоспециализированная клетка (нервная или эпителиальная), способное воспринять, трансформировать и передать энергию внешнего стимула в нервную систему. Понятие «рецептор» имеет и еще одно значение: мембранная белковая молекула, чувствительная к какому-либо стимулу; следует различать «клеточный рецептор» и «молекулярный рецептор». Далее мы будем использовать данный термин в первом значении.
Все рецепторы подразделяются на первично- и вторично-чувствующие (таблица 2, рис. 36). У первично-чувствующих рецепторов стимул воспринимается непосредственно (первично) сенсорным нейроном, тогда как в случае вторично-чувствующих рецепторов стимул воспринимается специализированной клеткой (обычно эпителиальной природы), которая затем передает сигнал чувствительному нейрону.
При воздействии адекватного стимула в рецепторах возникают особые электрические сигналы – рецепторные потенциалы, которые кодируют интенсивность и длительность воздействия стимула. Таким образом, любой вид сенсорного восприятия сопровождается трансдукцией — процессом перехода энергии любого стимула в электрический сигнал нервной системы. В таблице 2 представлено сравнение свойств различных сенсорных систем.
Рецепторный потенциал возникает в результате активации специальных рецепторных белковых молекул, чувствительных к каким-либо воздействиям.
Большинство рецепторных потенциалов являются возбуждающими, то есть проявляются в виде деполяризации мембраны. Они связаны с неселективным увеличением проницаемости для небольших катионов (натрий, калий, кальций). Вместе с тем, в сетчатке глаза и волосковых клетках уха были найдены и тормозные рецепторные потенциалы, которые гиперполяризуют мембрану.
Таблица 2. Свойства чувствительных систем организма
| Сенсорная система | Модальность | Энергия стимула | Класс рецепторов | Клеточный тип рецепторов | Вид Рецепторов |
| Зрительная | Зрение | Свет | Фоторецептор | Палочки и колбочки | Вторично-чувствующие |
| Слуховая | Слух | Звук | Механорецептор | Волосковые клетки (улитка уха) | Вторично- чувствующие |
| Вестибуляр-ная | Баланс | Гравитация | Механорецептор | Волосковые клетки (вестибулярный лабиринт), | Вторично- чувствующие |
| Соматосен-сорная | Прикосновение Проприоцепция Температурная чувствительность Боль | Давление Смещение Тепловая Химическая, тепловая, механическая | Механорецептор Механорецептор Терморецептор Хеморецептор, Терморецептор, Механорецептор | Кожные механорецепторы Мышечные и суставные рецепторы Тепловые и холодовые рецепторы Полимодальные, тепловые и механические ноцицепторы | Первично- чувствующие Первично- чувствующие Первично- чувствующие Первично- чувствующие |
| Вкусовая | Вкус | Химическая | Хеморецептор | Вкусовые сосочки | Вторично-чувствующие |
| Обонятель-ная | Запах | Химическая | Хеморецептор | Обонятельные нейроны | Первично- чувствующие |
Впервые рецепторные потенциалы были зарегистрированы Эдгаром Эдрианом на мышечных веретенах кожно-грудинной мышцы лягушки. В 1932 году за это и другие открытия в области физиологии нервной системы он был удостоен Нобелевской премии (рис. 35).
Возникновение рецепторного потенциала является начальным этапом сенсорного восприятия. Будучи локальным ответом, рецепторный потенциал затухает вблизи места возникновения, поэтому для успешной передачи сенсорного сигнала в нервной системе необходимо возникновение потенциала действия. Процесс передачи сенсорного сигнала в первично- и вторично-чувствующих рецепторах различается (рис. 36). В случае первично-чувствующих рецепторов на мембране дендрита сенсорного нейрона возникает рецепторный потенциал, и при достижении порогового уровня генерируется потенциал действия. Далее он проводится по дендриту, телу и аксону нейрона, достигая нервных окончаний аксона и вызывая выброс медиатора из них. В случае вторично-чувствующих рецепторов стимул воспринимается мембраной специализированной воспринимающей клетки, которая образует химический синапс с чувствительным нейроном. Рецепторный потенциал воспринимающей
Рис. 36. Принципиальные различия между первично-чувствующими и вторично-чувствующими рецепторами
В случае первично-чувствующих рецепторов (А) рецепторный потенциал (РП) возникает в окончании дендрита чувствительного нейрона. Затем происходит генерация потенциала действия (ПД), который проводится до тела клетки и далее по аксону.
В случае вторично-чувствующих рецепторов (Б) рецепторный потенциал возникает в специализированной рецепторной клетке, которая образует синапс с дендритом нейрона. Под действием рецепторного потенциала происходит изменение уровня секреции нейромедиатора из рецепторной клетки и возникновение генераторного потенциала (ГП) в дендрите чувствительного нейрона. Далее происходит генерация потенциала действия и его проведение к телу и далее по аксону чувствительного нейрона. Следует помнить, при некоторых видах рецепции (например, фоторецепции) дендрит чувствительного нейрона очень короткий, и проведение возбуждения по нему происходит электротонически, а потенциал действия генерируется в аксонном холмике.
клетки вызывает выброс медиатора, что приводит к появлению постсинаптического потенциала сенсорного нейрона, называемого также генераторным потенциалом. При достижении критического уровня деполяризации в нейроне возникает потенциал действия, который проводится в центральную нервную систему. Необходимо заметить, что гиперполяризационные рецепторные потенциалы, встречающиеся в фоторецепторах и волосковых клетках, уменьшают выброс медиатора, тем самым снижая величину мембранного потенциала и блокируя возникновение потенциалов действия в нейроне.
Более детально рассмотрим процесс трансдукции стимулов в различных видах рецепторов.
Дата добавления: 2015-07-04; просмотров: 3242; Опубликованный материал нарушает авторские права? | Защита персональных данных
Не нашли то, что искали? Воспользуйтесь поиском:
Лучшие изречения: Учись учиться, не учась! 11017 — | 8218 — или читать все…
Читайте также:
Источник
Фоторецепторы (палочки и колбочки) имеют два сегмента — внешний и внутренний (рис. 7.18). Во внутреннем сегменте расположены ядро, митохондрии, синаптические везикулы. Во внешнем — находятся столбики мембранных дисков или мешочков. В колбочках мешочки внешних сегментов образованные складками плазмолеммы, а в палочках мембранные диски отделены от мембраны клетки. Отработанные мембранные диски наружных сегментов с верхушки отделяются и фагоцитируются пигментными клетка

РИС. 7.18. Схема строения фоторецепторов: палочки и колбочки
мы, на их месте происходит формирование новых дисков.
Мембранные диски (мешочки) содержат зрительный пигмент родопсин , в состав которого входит белок — опсин и светопоглощающего аналог витамина А (ретинол), которая находится в форме 11-цис-ретиналя в темноте, когда свет не попадает на фоторецепторы. От аминокислотного состава опсина зависит длина световых волн, поглощает зрительный пигмент.
Палочки содержат только один тип опсина, который называется скотопсина, и максимальную чувствительность к свету с длиной волны 505 нм. В мембранных дисках палочек родопсин составляет до 90 % белков этих мембран.
Колбочки имеют три типа опсина зависимости от спектра поглощения света — голубого , зеленого , красного соответствии с длиной волны 440, 535, 565 нм, поэтому есть три разновидности колбочек, которые обеспечивают цветное зрение.
В темноте палочки деполяризована, их мембранный потенциал (МП) составляет около -40 мВ. Причиной деполяризации является высокая проводимость мембраны наружного сегмента фоторецепторов для ионов Na4 (рис. 7.19)
Ионы Na + входят в клетку через Na + -каналы мембраны наружного сегмента и транспортируются наружу Na + -K + -насоса мембраны внутреннего сегмента фоторецепторов. Na + -каналы поддерживаются открытыми благодаря циклическому гуанозинмонофосфата (цГМФ). Циклический гуанозинмонофосфат образуется из гуанозинтрифосфат (ГТФ) с помощью фермента гуанилатциклазы и связывается с Na + -каналов.

РИС. 7.19. Влияние света на транспорт ионов Na + и K + в фоторецепторных клетках сетчатки:
А — деполяризация; Б — гиперполяризация
Значительное количество митохондрий во внутреннем сегменте фоторецепторов обеспечивает образование АТФ, который необходим для работы Na + -K + -Hacocid в мембране внутреннего сегмента фоторецептора. Низкий уровень мембранного потенциала способствует высвобождению нейромедиатора глутамата в синаптическую щель (рис. 7.20).
Когда свет падает на фоторецепторы, в них возникает гиперполяризация . Поглощение кванта света родопсином начинает серию реакций, следствием которых является гидролиз циклического гуанозинмонофосфата (цГМФ) и закрытия натриевых каналов в наружном сегменте фоторецептора (рис. 7.21).
Формирование рецепторного гиперполяризацийного потенциала имеет такие шаги:
■ когда свет падает на сетчатку, ретиналь родопсина переходит из формы 11-цис-ретиналь в полный транс-ретиналь , благодаря чему родопсин через серию промежуточных веществ превращается в мета родопсин II ;
■ метародопсин II активирует на мембране дисков G-белок под названием трансдуцин , который в свою очередь активирует фермент фосфодиэстеразу ,
■ фосфодиэстеразы катализирует превращение циклического гуанозинмонофосфата ( ЦГ МФ ) до 5-ГМФ , поэтому концентрация цГМФ уменьшается;
■ уменьшение концентрации цГМФ приводит к закрыванию натриевых каналов , следствием чего является возникновение гиперполяризации в фоторецепторах.
Гиперполяризация мембраны фоторецептора приводит к уменьшению выделения тормозного медиатора, благодаря чему возникает деполяризация биполярных клеток сетчатки, информация от них передается на ганглионарные клетки, аксоны которых образуют зрительный нерв, в нервных волокнах которого генерируются ПД.
Свет в фоторецепторах вызывает каскад реакций, снижающих концентрацию внутриклеточного цГМФ и приводят к закрытию натриевых каналов, в результате чего уменьшается содержание в фоторецепторе не только Na +, но и Са2. В результате снижения концентрации Са 2+ активируется фермент гуанилотциклаза, что стимулирует синтез цГМФ и увеличение его содержания в клетке. Это приводит к торможению функции активированной светом фосфодиэстеразы. Оба эти процессы способствуют повышению содержания цГМФ и торможению активности фосфодиэстеразы, возвращает фоторецептор в исходное состояние открытия натриевых каналов.

РИС. 7.20. Механизм деполяризации мембраны наружного сегмента палочки в темноте

РИС. 7.21. Механизм гиперполяризации мембраны наружного сегмента палочки под воздействием света
Каждая ганглионарная клетка получает информацию от группы колбочек, которые образуют рецептивное поле. Рецептивные поля сетчатки расположены в виде круга, диаметр которого в центральной ямке может быть около 10 цм.
Колбочки в центре такого круга передают информацию через биполярные клетки непосредственно на ганглионарные клетки. Колбочки, расположенные на периферии круга, передают информацию опосредованно через горизонтальные клетки на биполярные и ганглионарные клетки.
Таким образом, в центре рецептивного поля осуществляется непосредственная активация биполярных и ганглионарных клеток ( «центральноувимкнена» клетка), так световой стимул активирует колбочки в центре поля и тормозит их на периферии через горизонтальные клетки ( «центральновимкнена» клетка) или наоборот (рис. 7.22) .
Организация рецептивного поля сетчатки обеспечивает передачу информации о контраст и интенсивность зрительных образов. Ощущение контраста формируется благодаря самой сигнализации в ганглионарной клетках, рецептивное поле которых лежит на границе между двумя уровнями освещения.

РИС. 7.22. Организация рецептивного поля и механизмы взаимодействия его центра с периферией
Интенсивность освещения кодируется разной частотой импульсов, возникающих в ганглионарной клетках при разном освещении фоторецепторов. При малом освещении информация поступает через биполярные клетки на ганглионарные только от палочек, которые имеют низкие пороги чувствительности к свету.
В сетчатке содержится много различных нейромедиаторов, в том числе возбуждающие — глутамат, ацетилхолин, дофамин, серотонин, различные нейропептиды, и тормозные нейромедиаторы — гамма-аминомасляная кислота (ГАМК), глицин. Амакринни клетки единственные в сетчатке, синтезирующих ацетилхолин, а горизонтальные — высвобождают ГАМК.
Формирование изображения на сетчатке фактически образуется тремя механизмами:
Первый — изображение возникает в результате действия света на фоторецепторы.
Второй — изображение образуется благодаря передаче информации нейромедиаторами на биполярные клетки в центре действия световых лучей, и при участии горизонтальных клеток — на периферии круга рецептивного поля.
Третий — осуществляется при участии ганглионарной и амакринних клеток, которые модифицируют изображения на периферии круга рецептивного поля. Изображение почти не модифицируется в зрительных центрах — латеральных коленчатых телах — и информация о нем доходит до затылочной сенсорной зоны коры.
Источник
Рецепторный потенциал. Рецепторный потенциал тельца Пачини
Все сенсорные рецепторы имеют одну общую особенность. Независимо от типа стимула, возбуждающего рецептор, непосредственным результатом действия стимула является изменение мембранного потенциала рецептора. Это изменение потенциала называют рецепторным потенциалом.
Механизмы развития рецепторных потенциалов. В разных рецепторах развитие рецепторного потенциала может быть вызвано одним из нескольких способов: (1) механической деформацией рецептора, ведущей к растяжению его мембраны и открытию ионных каналов; (2) действием химического вещества на мембрану, в результате также открываются ионные каналы; (3) изменением температуры в области мембраны рецептора, что меняет ее проницаемость; (4) влиянием электромагнитного излучения, например действием света на рецепторы сетчатки глаза, прямо или косвенно изменяющим свойства ионных каналов рецепторной мембраны.
В дальнейшем будет показано, что эти четыре способа возбуждения рецепторов в целом охватывают все известные типы сенсорных рецепторов. Во всех случаях основной причиной изменения мембранного потенциала является изменение проницаемости мембраны рецептора для ионов, которые в результате более или менее легко диффундируют через мембрану, изменяя трансмембранный потенциал.
Максимальная амплитуда рецепторного потенциала. Для большинства сенсорных рецепторов максимальная амплитуда их рецепторных потенциалов составляет около 100 мВ, но такого уровня рецеп-торный потенциал достигает только при чрезвычайной интенсивности сенсорного стимула.
Известно, что эта же величина мембранного потенциала регистрируется при развитии потенциала действия и соответствует состоянию максимальной проницаемости мембраны для ионов Na+.
Отношение между рецепторным потенциалом и потенциалами действия. Как показано на рисунке, когда амплитуда рецепторного потенциала поднимается выше порога возбуждения нервного волокна, связанного с рецептором, в волокне возникают потенциалы действия. При этом чем выше ре-цепторный потенциал порогового уровня, тем выше частота потенциалов действия.
Рецепторный потенциал тельца Пачини
Здесь следует вспомнить анатомическую структуру тельца Пачини, показанного на рисунке. Видно, что тельце имеет центральное нервное волокно, проходящее через его сердцевину. Волокно окружено капсулой, состоящей из многочисленных концентрических слоев, поэтому давление в любой точке с внешней стороны тельца растягивает или каким-либо другим образом деформирует центральное волокно.
На рисунке показано центральное волокно тельца Пачини после удаления всех слоев капсулы, кроме одного. Концевая часть центрального волокна, расположенная внутри капсулы, не покрыта миелином, но непосредственно перед выходом из капсулы волокно становится миелинизированным (миелиновая оболочка показана синим), и таким оно входит в состав периферического чувствительного нерва.
На рисунке также показан механизм развития рецепторного потенциала в тельце Пачини. Обратите внимание на небольшую область концевой части волокна, деформированную давлением на тельце. В ответ на деформацию в мембране открылись ионные каналы, через которые положительно заряженные ионы Na+ диффундируют внутрь волокна. Это сдвигает мембранный потенциал волокна в положительную сторону, что и является рецепторным потенциалом.
Его развитие, в свою очередь, вызывает появление локального кругового тока, распространяющегося вдоль нервного волокна (на рисунке ток обозначен стрелками). Когда этот местный ток деполяризует мембрану волокна в области первого перехвата Ранвье, расположенного внутри капсулы тельца Пачини, генерируются типичные потенциалы действия, которые проводятся вдоль нервного волокна к центральной нервной системе.
Отношение между интенсивностью стимула и рецепториым потенциалом. На рисунке показано изменение амплитуды рецепторного потенциала, которое отмечается в условиях эксперимента при постепенном усилении механического давления (увеличения силы стимула) в области сердцевины тельца Пачини. Видно, что сначала амплитуда быстро возрастает, но постепенно, по мере нарастания силы стимула, прирост рецепторного потенциала замедляется.
В свою очередь, почти пропорционально увеличению амплитуды рецепторного потенциала возрастает частота потенциалов действия, передаваемых от сенсорных рецепторов. Сопоставив эту закономерность с данными, представленными на рисунке, можно понять, что при очень интенсивном раздражении рецептора прирост частоты потенциалов действия постепенно снижается. Это чрезвычайно важный принцип, применимый почти ко всем сенсорным рецепторам.
Он позволяет рецептору быть чувствительным к очень слабому сенсорному раздражению и все же не достигать максимальной активности до тех пор, пока сенсорное раздражение не станет чрезмерным. Благодаря этому свойству диапазон реакции рецептора исключительно велик: от очень слабой активности до очень интенсивной.
— Также рекомендуем «Адаптация рецепторов. Механизмы адаптации рецепторов»
Оглавление темы «Передача нервного сигнала. Сенсорная нервная система»:
1. Рецепторный потенциал. Рецепторный потенциал тельца Пачини
2. Адаптация рецепторов. Механизмы адаптации рецепторов
3. Быстроадаптирующиеся рецепторы. Нервные волокна
4. Передача нервный сигналов. Пространственная суммация нервных импульсов
5. Механизмы передачи нервных сигналов. Пороговые и подпороговые нервные стимулы
6. Дивергенция нервных сигналов. Конвергенция нервных сигналов
7. Последействие нервного сигнала. Синоптическое последействие нервного импульса
8. Импульсная активность нервных контуров. Ритмичная активность нервных контуров
9. Тормозные контуры нервной системы. Синоптическое утомление
10. Соматосенсорная система. Тактильные ощущения
Источник

