
Пигменты рецепторов сетчатки глаза
Зрительная фототрансдукция представляет собой комплекс процессов, который отвечает за изменение (фототрансформацию) пигментов и последующую их регенерацию. Необходимо это для передачи информации из внешнего мира к нейронам. Благодаря биохимическим процессам, при влиянии света с различной длиной волны, возникают структурные изменения в строении пигментов, которые находятся в бислойном липидном участке мембран внешней доли фоторецептора.

Изменения в фоторецепторах
Фоторецепторы всех позвоночных животных, включая человека, могут реагировать на световые лучи путем изменения фотопигментов, которые располагаются в бислойных мембранах в области внешней доли колбочек и палочек.
Сам зрительный пигмент представляет собой белок (опсин), который является производным витамина А. Сам бета-каротин содержится в пищевых продуктах, а также синтезируется в клетказ сетчатки (фоторецептоный слой). Эти опсины ил хромофоры в связанном состоянии локализуются в глубине биполярных дисков в зоне внешних долей фоторецепторов.
Около половины опсинов приходится на бислойный липидный слой, который связан снаружи короткими петлями белка. Каждая молекула родопсина имеет в своем составе семь трасмембранных участков, которые окружают хромофор в бислое. Хромофор располагается горизонтально в мембране фоторецептора. Внешний диск мембранного участка имеет большое количество зрительных молекул пигмента. После того, как был поглощен фотон света, вещество пигмента переходит из одной изоформы в другую. В результате этого молекула претерпевает конформационные изменения, а структура рецептора восстанавливается. При этом метародопсин активирует G-белок, что запускает каскад биохимических реакций.
Фотоны света воздействуют на зрительный пигмент, что приводит к активации каскада реакций: фотон – родопсин – метародопсин – трансдуцин – фермент, который гидролизует цГМФ.В результате этого каскада формируется закрывающаяся мембрана на внешнем рецепторе, которая связана с цГМФ и отвечает за работу катионного канала.
В темноте через открытые каналы проникают катионы (в основном ионы натрия), которые приводят к частичной деполяризации ячейки фоторецептора. При этом этот фоторецептор выбрасывает медиатор (глутамат аминокислоты), который воздействует на инаптические окончания нейронов второго порядка. При незначательном световом возбуждении молекула родопсина изомеризуется в активную форму. Это приводит к закрытию ионного трансмембранного канала, и, соответственно, останавливает катионный поток. В результате клетка фоторецептора гиперполяризуется, а медиаторы перестают выделяться в зоне контакта с нейронами второго порядка.
В темноте через трансммбранные каналы осуществляется поток ионов натрия (80%), кальция (15%), магния и других катионов. Чтобы удалить избыток кальция и натрия во время темноты, в клетках фоторецепторов действует катионный обменник. Ранее считалось, что кальций участвует в фотоизомерации родопсина. Однако в настоящее время получены доказательства того, что этот ион играет и другие роли в фототрансдукции. За счет присутствия достаточной концентрации кальция, палочковые фоторецепторы становятся более восприимчивыми к свету, а также значительно увеличивается восстановление этих клеток после освещения.
Колбочковые фоторецепторы способны приспособиться к уровню освещения, поэтому человеческий глаз способен воспринимать объекты при разном освещении (начиная от теней под деревом и заканчивая предметов, расположенных на блестящем освещенном снегу). Палочковые фоторецепторы имеют меньшую приспособляемость к уровню освещения (7-9 единиц и 2 единицы для колбочек и палочек, соответственно).
Фотопигменты экстерорецепторов колбочек и палочек сетчатки глаза
К фотопигментам колбочкового и палочкового аппарата глаза относят:
- Йодопсин;
- Родопсин;
- Цианолаб.
Все эти пигменты отличаются друг от друга аминокислотами, которые входят в состав молекулы. В связи с этим пигменты поглощают определенную длину волны, точнее диапазон длин.
Фотопигменты экстерорецепторов колбочек
В колбочках сетчатки глаза располагается йодопсин и разновидность йодопсина (цианолаб). Все выделяют три типа йодопсина, которые настроены на длину волны в 560 нм (красный), 530 нм (зеленый) и 420 нм (синий).
О существовании и идентификации цианолаба
Цианолаб представляет собой разновидность йодопсина. В сетчатке глаза синие колбочки располагаются регулярно в периферической зоне, зеленые и красные колбочки локализуются хаотично по всей поверхности сетчатки. При этом плотность распределения колбочек с зеленым пигментов больше, чем красных. Наименьшая плотность отмечается у синих колбочек.
В пользу теории трихромазии свидетельствуют следующие факты:
- Была определена спектральная чувствительность двух пигментов колбочки при помощи денситометрией.
- С использованием микроспектрометрии было определено три пигмента колбочкового аппарата.
- Был идентифицирован генетический код, ответственный за синтез красных, синих и зеленых колбочек.
- Ученым удалось изолировать колбочки и измерить их физиологический ответ на облучение светом с определенной длинной волны.
Теория трохромазии раньше была не в состоянии объяснить наличие четырех основных цветов (синий, желтый, красный, зеленый). Также было затруднительно объяснить, почему люди-дихроматы способны различать белый и желтый цвета. В настоящее время открыт новый фоторецептор сетчатки, в котором роль пигмента исполняет меланопсин. Это открытие расставило все по местам и помогло ответить на многие вопросы.
Также в недавних исследованиях при помощи флуоресцентного микроскопа были изучены срезы сетчатки птиц. При этом было выявлено четыре типа колбочек (фиолетовая, зеленая, красная и синяя). За счет оппонентного цветного зрения фоторецепторы и нейроны дополняют друг друга.
Фотопигмент палочек родопсин
Родопсин относится к семейству G-связанных белков, который так назван из-за механизма трансмембранной передачи сигнала. При этом в процесс вовлекаются G-белки, расположенные в примембранном пространстве. При исследовании родопсина была установлена структура этого пигмента. Это открытие очень важно для биологии и медицины, потому что родопсин является родоначальником в семействе GPCR-рецепторов. В связи с этим его строение используется в изучении всех остальных рецепторов, а также определяет функциональные возможности. Родопсин назван так, потому что имеет ярко-красную окраску (с греческого он дословно переводится как розовое зрение).

Дневное и ночное зрение
Изучая спектры поглощения родопсина, можно заметить, что восстановленный родопсин отвечает за восприятие света в условиях низкой освещенности. При дневном свете этот пигмент разлагается, и максимальная чувствительность родопсина смещается в синюю спектральную область. Это явление получило название эффект Пуркинье.
При ярком освещении палочка перестает воспринимать дневные лучи, а эту роль на себя берет колбочка. При этом происходит возбуждение фоторецепторов в трех областях спектра (синий, зеленый, красный). Далее эти сигналы преобразуются и направляются в центральные структуры мозга. В результате формируется цветное оптическое изображение. Для полного восстановления родопсина в условиях низкой освещенности требуется коло получаса. В течение всего этого времени происходит улучшение сумеречного зрения, которое достигает максимума по окончании периода восстановления пигмента.
Биохимик М.А. Островский провел ряд фундаментальных исследований и показал, что палочки, содержащие пигмент родопсин, участвуют в восприятии объектов в условиях низкого освещения и отвечают за ночное зрение, которое имеет черно-белую окраску.
Источник
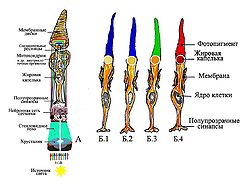
Рис.1a. Принципиальная схема организации цветного зрения (на примере сетчатки цыплёнка).
А. Фоторецептор-колбочка с пигментами. Свет проходит сквозь хрусталик и фиксируется (фильтруется) соответствующим пигментом в конусной мембране колбочки, расположенной на её «дне». Как видим имеются четыре сечения мембраны с пигментами четырёх цветов. (У человека три сечения цветов пигмента RGB — Трихроматизм (цветное зрение). Колбочка А с четырьмя вариантами работы при её освещении.
Б.1-Б.4 У птиц обычно четыре варианта работы колбочки (они «тетрахроматы»), что позволяют им синтезировать многоцветное изображение, и различать цвета лучше, чем человек. Обработка полученных разностных сигналов, происходящая в нейронной сети сетчатки (десятки типов клеток) обеспечивает возможность ясного различения тысяч цветов и оттенков. Важно, что схема цветного зрения человека может быть показана в виде колбочки A, способная принимать любой луч основного цвета RGB в трёх соответственных сечениях внешней мембраны S,M,L — трихроматизма.[1]
Рис. 2a. В Диаграмме CIE rg цветность пространства показывает построенный треугольник, определяющий цветовое пространство CIE XYZ (См. XYZ (цветовая модель)).
Треугольник Cb-Cg-Cr это только xy=(0,0), (0,1), (1,0), треугольник CIE xy— цветность пространства. Линия, соединяющая Cb и Cr это линия нулевой яркости на цветовом графике (alychne).
Обратите внимание, что спектральный локус проходит через rg=(0,0) на 435.8 нм, через rg=(0,1) в 546.1 нм и через rg=(1,0) при 700 нм. (По старой системе применялось цветовое пространство RGB и рассчитывалось без учёта линейной функции, а область полученной цветовой палитры была внутри треугольника с углами в точках 430 нм, 540 нм и 570 нм).
Значение же энергии самой яркой точки (E) при rg=xy=(1/3,1/3) равное (белый цвет). (См. также XYZ (цветовая модель), Цветовые координаты)[2]
Зрительные пигменты и передача зрительного сигнала (зрительная фототрансдукция (версия Миг)) — комплекс понятий для описания фототрансформации пигментов и их регенерации; процессов передачи сигнала, происходящих в глазе позвоночных животных. Эти биохимические процессы проходят при воздействии света с различной длиной волны (разного цвета), связанные с изменениями в структуре и взаимодействиях зрительных пигментов, находящихся в бислойных биомембранах, во внешней доле фоторецептора (в мембране колбочек, палочек).
Изменения в фоторецепторах[править]
Фоторецепторы позвоночных животных, например, приматов, человека реагируют на свет (цвет) посредством изменений в содержащихся в них зрительных фотопигментов (см.Опсины (версия Миг). Зрительные пигменты фоторецепторов сетчатки глаза и передача зрительного сигнала) происходит в бислойных мембранах внешней доли фоторецептора (колбочки, палочки).
Зрительный пигмент состоит из белка, названного опсином хромофора — производного витамина А, известного как вещество, относящееся к сетчатке глаза. Витамин А — производное бета-каротина (cм. также G-белки), находящегося в нашей пище, белки синтезируется в клетке фоторецептора (см. выше) и описываются под общим названием опсины и хромофоры, они в связанном виде находятся в глубине внешних долей биполярных дисков мембран фоторецепторов (колбочек, палочек).
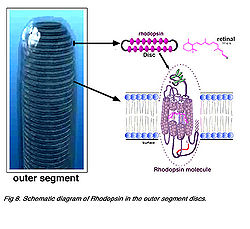

- Рис. 8. Схематическая диаграмма родопсина во внешних дисках доли;
- Рис. 9. Структурная модель родопсина.
Приблизительно 50 % опсинов находится в пределах бислойной липидной мембраны, связанной короткими петлями белка снаружи. Каждая молекула rhodopsinа состоит из семи этих трансмембранных частей, окружающих хромофор (он относится к сетчатке глаза 11 СНГ) в двойном слое липида. Хромофор очевидно находится горизонтально в мембране и связан в остатке лизина спирали семь (Hargrave и др. 1984, Hargrave и McDowell, 1992). Каждый внешний диск доли мембраны, конечно, содержит много (тысячи) визуальных молекул пигмента. После поглощения фотона света, то относящийся к сетчатке глаза пигмент изомеризуются из формы с 11 СНГ к форме после произшедших всех преобразований, которые начинают конформационные изменения в молекуле, что приводит к восстановлению структуры рецептора. Несколько посредников сформированы при отбеливании метародопсина II, который активизирует G-белок transducina и создаёт дальнейший каскад событий, описанных ниже (см. обзор Hargrave и McDowell (1992) и Стрельцом, 1995).
Свет преобразует зрительный пигмент через следующий ферментный каскад: фотоны — родопсин — активизированный rhodopsin (metarhodopsin II) — GTP обязательный белок (трансдуцин) — фермент, гидролизирующий cGMP (cGMP-phosphodiesterase). В итоге возникает закрывающаяся мембрана экстерорецептора, связавшая cGMP с регулируемым катионным каналом.
В темноте устойчивый поток течет в открытые каналы, которые несут главным образом ионы Na, составляя «темный поток», который частично деполяризует ячейку фоторецептора. Таким образом, деполяризованный фоторецептор выпускает медиатор (предполагая, что это является глутаматом аминокислоты) из его синаптических терминалов на нейроны второго заказа в темноте. На легком (световом) возбуждении молекулы родопсина приводят isomerized к активной форме вышеупомянутого следующего каскада, приводя к закрытию каналов катиона мембраны фоторецептора, останавливая темновой поток и заставляя мембранные клетки фоторецептора гиперполяризовывать и останавливать выпуск медиатора к нейронам второго заказа (см. Stryer, 1991; Yau, 1994, и Kawamura, 1995, для обзоров.[3]
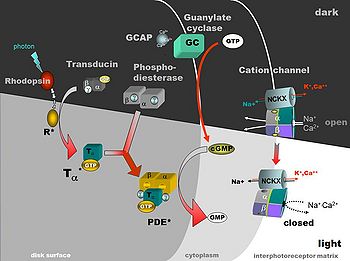
Рис. 10. Активация Rhodopsin при свете и каскаде фототрансдукции.
Составляющими «Темного потока» являются главным образом: приток Na + компонентов (80 %), Ca2 + компонентов — (15 %) и Mg2 + компонентов — (5 %) (Yau, 1994). В темноте необходим механизм, чтобы удалить Ca2 + так же как лишний Na +, и это, как полагают, сделано через теплообменник натрия/кальция в мембранах внешней доли фоторецептора. Ca2 +, как когда-то полагали, предназначен, чтобы быть вторым посыльным в соединении rhodopsin фотоизомеризации к мембранным событиям, но как теперь доказано имеет вторичную, но важную регулирующую роль в фототрансдукции. Хотя это непосредственно не участвует в каскаде трансдукции, но это действительно улучшает сигнальную способность палочек в ускорении восстановления после освещения и регулирует падение чувствительности палочек в устойчивом освещении (Yau, 1994). Последний открытый эффект — это механизм для легкой адаптации.
Фоторецепторы колбочки могут приспособиться к уровню освещения так, что наша зрительная система может видеть в диапазоне от плотных теней под деревом, до объектов на ярком снегу солнечного света при изменении интенсивности в пределах 7‒9 единиц регистрации легкой интенсивности (Normann и др., 1991). Фоторецепторы же палочек приспосабливаются в диапазоне 2 единиц регистрации второстепенной интенсивности и система приспособления объединена с адаптацией сети через целую визуальную систему, и позволяет целым 5 единицам регистрации второстепенной адаптации интенсивности в палочке, которая открывает зрение (Yau, 1994).
Фотопигменты экстерорецепторов колбочек и палочек сетчатки глаза[править]
Разные опсины (версия Миг) различаются аминокислотами в составе молекул, и поглощают свет в несколько различном диапазоне длин волн, как ретиналь-связанные молекулы.
Фотопигменты экстерорецепторов сетчатки глаза колбочек[править]
У человека идентифицировано три вида фотопигментов колбочек йодопсина (фотопсина):
- 560 nm: rot-sensitives L‒Iodopsin mit L-Photopsin, Шаблон:UniProt. Defekte am OPN1LW-Gen sind die Ursache der Protanopie (красный цвет).
- 530 nm: grün-sensitives M‒Iodopsin mit M-Photopsin, Шаблон:UniProt. Defekte am OPN1MW-Gen sind die Ursache der Deuteranopie (зелёный цвет).
- 420 nm: blau-sensitives S-Iodopsin mit S-Photopsin, Шаблон:UniProt. Defekte am OPN1SW-Gen sind die Ursache der Tritanopie (синий, УФ цвет).
| Cone type | Name | Range | Peak wavelength[4][5] |
|---|---|---|---|
| S (OPN1SW) — «tritan», «cyanolabe» | β | 400—500 nm | 420‒440 nm (УФ, синий) |
| M (OPN1MW) — «deutan», «chlorolabe» | γ | 450—630 nm | 534‒545 nm (зелёный) |
| L (OPN1LW) — «protan», «erythrolabe» | ρ | 500—700 nm | 564‒580 nm (красный) |
О существовании и идентификации цианолаба[править]
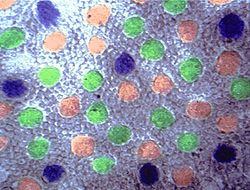
Рис. S. Распределение рецепторов в сетчатке бабуина . Синие колбочки были распределены регулярно в периферии, красные и зеленые колбочки были распределены беспорядочно всюду. Плотность распределения зелёных колбочек больше, чем красных, больше чем синих. Данные получены доктором Марком и его лабораторией впервые, где видны синие колбочки с предполагемым фотопигментом цианолабом.[6]
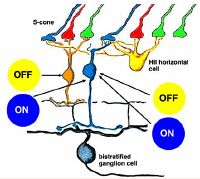
Рис.9. Нейроны троп Булочки — «синей» Колбочки-S, расположенной в периферийной зоне ямки фовеа, воспринимающей сфокусиованные синие лучи света предметной точки на колбочки M/L в центральной ямке фовеа 0,2 мм из блока основных лучей RGB сфокусированного луча. Оппонентно выделенные биосигналы синего цвета ON в системе отбора цвета из пары синий-жёлтый с участием горизонтальных клеток H, что говорит о наличии не выделеннрго пока фотопигмента синего цвета цианолаба. При этом также OFF желтый сигнал может быть предоставлен по центру диффузному биполярному типу клеток Bi. (См. более подробно также в Тропы синих колбочек-S сетчатки глаза) [7]
* С точки зрения принципа трихроматизма следует:
- Идентификация спектральной чувствительности двух пигментов колбочки, относящейся к сетчатке глаза, денситометрией Руштона (Руштон, 1963) (См. Денситометрия (версия Миг), Денситометр (версия Миг)).
- Идентификация трех пигментов колбочки микроспектрометрией (Марки, Dobelle и MacNichol, 1964).
- Инентификация генетического кода для красных колбочек — L, зелёных колбочек — М. и синих колбочек — S (Nathans и другие, 1986a, b).
- Цвет, соответствующий функциям.
- Изолируя фоторецепторы и измерение их физиологического repsonses как функция длины волны (Baylor и другие, 1984).
- Спектральные размеры чувствительности (Wald-Marre — фунуционирование спектральной чувствительности и функционирование «(~pidots) Турникетов» механизмов).
Если теория трихроматизм (trichomatic) ранее была не в состоянии составлять четыре уникальных цвета: красный, зеленый, желтый и синий, и также не в состоянии объяснить, почему dichromats может чувствовать белый и желтый, то в настоящее время Трихроматизм с открытием третьего фоторецептора ipRGC сетчатки с фотопигментом меланопсином, с расшифровкой изменчивости опсинов, которые входят в мембраны клеток фоторецепторов палочек и колбочек в виде разновидностей опсина как родопсин (версия Миг) (палочки), йодопсин (версия Миг)(колбочки) и др., позволяют дифференциацироваться колбочкам и реагировать на нормализованные спектральные зоны лучей S,M,L (синих, зелёных, красных), участвуя в оппонентном отборе более ярких сигналов.
Недавние открытия ретиномоторной реакции фоторецепторов палочек и колбочек, проведенные флюоресцентные микроскопические исследования живых срезов сетчаток птиц дали возможность рассмотреть клетки колбочек, палочек на молекулярном уровне в цвете и стерео, где мозаика сетчатки содержит блоки из четырёх колбочек (четырехроматизм): фиолетовой, синей, зелёной и красной и т. д. Модель оппонентного цветного зрения в сочетании с работой нейронов мозга рассматривается на уровнях рецепторном и нейронном, которые дополняют друг друга. И вообще уже вопрос многокомпонентной модели цветного зрения как трихромаик, четырехроматик и т. д. не подлежит рассмотрению. (См. Цветное зрение у птиц (версия Миг), Лаборатория Р.Е.Марка (версия Миг), Ретиномоторная реакция фоторецепторов (версия Миг), Фоточувствительные клетки сетчатки ipRGC (версия Миг), Опсины (версия Миг)).
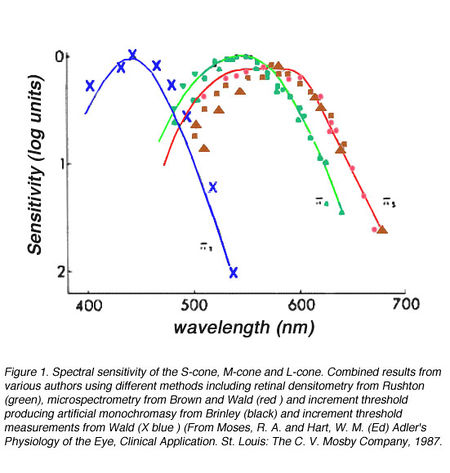
Рис.1. Спектральная чувствительность S-колбочек, М-колбочек и L-колбочек. Комбинированные результаты от разных авторов, используя различные способы, в том числе сетчатки денситометрия от Раштон (т и Ñ), microspectrometry от Коричневого и Вальд (n и «) и прирост порог производству искусственного monochromasy от Brinley (D и s) и прирост порог измерений от Wald (5) (От Моисея, р. а., Харт, в. м. (Ред.), Адлер » Физиология Глаза, Клиническое Применение. Сент-Луис: C. V. Мосби Компании, 1987 [8]
Получены снимки колбочек (см.рис. S), графики трёх колбочек (см.рис.1), воспринимающих основные лучи КЗС — S,M,L, на которых видна графика точек синих лучей S, выделяемых S-колбочками, также видны сами синие колбочки, снятые в плане фокальной поверхности сетчатки. Вопрос пока не открытого фотопигмента йодопсина цианолаба не означает, что нет синей колбочки, и что разновидности опсина не содержат пока не открытый синий фотопигмент опсинов — цианолаб, который находится в колбочках.
Фотопигмент палочек родопсин[править]
Родопсин принадлежит к обширному семейству G-белоксопряжённых рецепторов (GPCR-рецепторов), названному так за механизм трансмембранной передачи сигнала, основанный на взаимодействии с внутриклеточными примембранными G-белками. Появление его пространственной структуры, полученной с высоким экспериментальным разрешением, является очень важным событием для биологии и медицины, поскольку родопсин как «родоначальник» семейства А GPCR-рецепторов является своего рода «моделью» структуры и функций множества других рецепторов, чрезвычайно интересных с фундаментальной и практической (фармакологической) точек зрения. Свое название родопсин получил за ярко-красный цвет (по-гречески rhodon — означает «розовый», a opsis — зрение).
Дневное и ночное зрение[править]
Из спектров поглощения родопсина видно, что восстановленный родопсин (при слабом «сумеречном» освещении) отвечает за ночное зрение, а при дневном «цветовом зрении» (ярком освещении) разлагается и максимум его чувствительности смещается в синюю область. Это наглядно показывает известный Эффект Пуркинье (версия Миг)..[9]
При достаточном, дневном освещении освещении палочка автоматически уходит из зоны восприятия дневных лучей КЗС и для восприятия дневного света открываются колбочки (см. Ретиномоторная реакция фоторецепторов сетчатки глаза), которые выделяют основные лучи видимого спектра сфокусированной предметной точки. Сигналы этих лучей равны длинам волн S,M,L (синих, зелёных, красных) не в цвете). Посылая их в мозг, в зрительных отделах создаётся цветное оптическое изображение стерео. Полное восстановление родопсина у человека (в темноте или при слабом освещении) занимает около 30 минут; в течении всего этого периода чувствительность нашего «сумеречного зрения» постепенно увеличивается, достигая максимума.
На основе проведённых фундаментальных исследований академика биохимика М. А. Островского в области работы фотопигмента родопсин в палочках[10] (Биохимия зрения и свободно-радикальное окисление (версия Миг)) было также подтверждено, что палочки с фотопигментом родопсином участвуют в зрительном процессе в условиях сумеречного и ночного зрения (чёрно-белого).
См. также[править]
- Опсины (версия Миг)
- G-белки
- Эволюция и генетика фотопигментов сетчатки глаза
Примечания[править]
Источник

