
Обработка информации в сетчатке
В сетчатке происходит не только собственно фоторецепция (преобразование световых раздражителей в электрические сигналы), но и процессы первичной обработки зрительной информации, направленные на выделение значимых и удаление незначимых компонентов изображения, например, выделение контраста, выделение цветов и пр. Т. о., сетчатку можно сравнить с фотоаппаратом и системой компьютерной предобработки изображения.
Соответственно своим сложным функциям сетчатка имеет и сложное гистологическое строение. В ней выделяют клетки пяти основных типов:
— фоторецепторы – палочки и колбочки;
— биполярные клетки;
— ганглиозные клетки;
— горизонтальные клетки;
— амакриновые клетки.
Функции этих клеток следующие:
Фоторецепторы, биполярные и ганглиозные клетки отвечают за фоторецепцию и проведение зрительной информации: фоторецепторы преобразуют световой раздражитель в электрический сигнал, от них этот сигнал передается к биполярным клеткам и далее – к ганглиозным клеткам, аксоны которых покидают глазное яблоко, образуя зрительный нерв.
Горизонтальные и амакриновые клетки отвечают за первичную обработку зрительной информации, образуя вместе с остальными клетками различные нейронные контуры.
Конвергентный контур, рецептивные поля сетчатки и острота зрения.
На одной ганглиозной клетке конвергируют (через биполярные клетки) несколько фоторецепторов (в среднем – около 60). Степень этой конвергенции определяет величину рециптивного поля сетчатки, а тем самым – остроту зрения. При этом:
· В периферических отделах сетчатки, отвечающих за периферическое (боковое) зрение, эта конвергенция выражена гораздо больше (до 600 фоторецепторов на 1 ганглиозную клетку), что обеспечивает высокую чувствительность к слабому свету и позволяет быстро реагировать даже на небольшие изменения освещенности и движущиеся тени, однако не позволяет различать их детали (острота периферического зрения мала);
· В центральных отделах сетчатки – жёлтом пятне и особенно центральной ямке – эта конвергенция почти не выражена («один фоторецептор – одна ганглиозная клетка»), что позволяет различать мельчайшие детали объектов (острота центрального зрения велика).
Контур латерального торможения и выделение контраста.
Роль тормозных нейронов играют горизонтальные клетки.
Контур реципрокного торможения, выделение цветов и противоположные цвета.
Одна и та же ганглиозная клетка может возбуждаться при активации колбочек одного цвета и тормозиться – при возбуждении колбочек другого цвета. Такие реципрокные соотношения характерны для колбочек, спектры которых перекрываются:
· Колбочек красного цвета и зеленого цвета;
· Колбочек синего цвета, с одной стороны, и красного и зеленого цветов – с другой.
В связи с этим выделяют пары противоположных цветов: красный – зелёный; синий – жёлтый (сочетание красного и зелёного)
Выделение изменений.
Важнейшее свойство обработки информации в сетчатке заключается в том, что большинство ганглиозных клеток в конечном счёте реагируют не на постоянный уровень освещенности соответствующего рецептивного поля, а на его изменения. Благодаря этому мы реагируем на движущиеся объекты и изменения освещенности, но быстро перестаем воспринимать неподвижные неизменные объекты.
37. Теории цветоощущения. Основные формы нарушения цветового восприятия. Периметрия.
Цвет имеет три основных показателя: тон (оттенок), интенсивность и насыщение.
Теории цветоощущения.
Цветовосприятие – функция колбочек. Наибольшим признанием пользуется трехкомпонентная теория механизма восприятия цветов (теория Ломоносова – Юнга(Янг) — Гельмгольца). Согласно этой теории, в сетчатке глаза размещены три различных типа колбочек, из которых каждый обладает совершенно определенной спектральной чувствительностью. Одни чувствительны к красному цвету, другие- к зеленому, а третьи к синему(фиолетовому). Всякий цвет оказывает действие на все три цветоощущающих элемента, но в разной степени.
Эти возбуждения суммируются зрительными нейронами и, дойдя до коры дают ощущение того или иного цвета.
Согласно другой теории — теории опонентных цветов, предложенной Э.Герингом в ХIХ веке (Во многих сенсорных системах используют оппонентные пары: тепло/холод; черное/белое и т.д.)
Для доказательства использовался метод микроспектрофотометрических изменений одиночных колбочек. Геринг предположил, что имеются четыре основных цвета красный, желтый, зеленый и синий — и что они попарно связаны с помощью двух антагонистических механизмов-зелено-красного механизма и желто-синего механизма. Предполагается такой же механизм для ахроматических дополнительных цветов белого и черного. Из-за полярного характера восприятия этих цветов Геринг назвал эти цветовые пары «оппонентными цветами»
Из его теории следует, что не может быть таких цветов, как «зеленовато-красный» и » синевато-желтый». Таким образом теория оппонентных цветов постулирует наличие антагонистических цветоспецифических нейронных механизмов.
Если нейрон возбуждается под действием зеленого светового стимула, то красный стимул должен вызывать его торможение.
В настоящее время признаются обе теории- так как при использовании микроэлектродных отведений было доказано, что импульсы в ганглиозных клетках могут возникать в следующих случаях (при действии любого света (доминаторы) и при освещении только одни светом (модуляторы)
Установлено 7 типов модуляторов оптимально реагирующих на свет с разной длиной волны (от 400до 600 нм).
Различие цветов.
Фотоны различаются по длине волны: чем выше частота (и энергия) фотона, тем короче его волна. Поскольку разные вещества (гемоглобин, хлорофилл) испускают фотоны с разной длиной волны, способность различать световые лучи с разной длиной волны имеет важное приспособительное значение: очевидно, что живое существо, издалека видящее пятно хлорофилла (например, лес) или гемоглобина (кровь), имеет явные преимущества.
Человек различает разные длины волн в виде субъективного ощущения цвета; это называется цветовым зрением.
Самой большой длине волны (самой низкой частоте и энергии фотонов) соответствует красный цвет, самой короткой (наибольшей частоте и энергии фотонов) – фиолетовый. Цвета между красным и фиолетовым располагаются в известной последовательности красный – оранжевый – жёлтый – зелёный – голубой – синий – фиолетовый («Каждый Охотник Желает Знать Где Сидит Фазан»).
Наш глаз воспринимает длины волн только в диапазоне 400-700 нм. Фотоны с длиной волны выше 700 нм относятся к инфракрасному излучению, мы воспринимаем их в форме тепла. Фотоны с длиной волны ниже 400 нм относятся к ультрафиолетовому излучению, они из-за своей высокой энергии способны оказывать повреждающее действие на кожу и слизистые; после ультрафиолетового идёт уже рентгеновское и гамма-излучение.
Цветовое зрение возможно потому, что разные колбочки обладают разной спектральной чувствительностью, то есть разной чувствительностью к фотонам разной длины волны (это обусловлено тем, что в разных колбочках содержатся разные зрительные пигменты). Различают колбочки трёх видов:
· Колбочки синего цвета;
· Колбочки зелёного цвета;
· Колбочки красного цвета.
Цвет кодируется соотношением между возбужденными колбочками разного типа.
Источник
В фоторецепторах происходит взаимодействие
кванта света с пигментом. Рассмотрим
эти процессы на примере хорошо изученного
взаимодействия света с родопсином.
Родопсин представляет собой
высокомолекулярное соединение (мол.
масса 270000), состоящее из альдегида
витамина А — ретиналяи белкаопсина.
Фотохимические превращения зрительных
пигментов начинаются с поглощения ими
фотона и перехода на более высокий
энергетический уровень, сопровождающийся
ихстереоизомеризацией. При этом
происходит поэтапное образование ряда
промежуточных продуктов, в конечном
итоге чего разрывается связь ретиналя
с опсином (рис. 6.23).
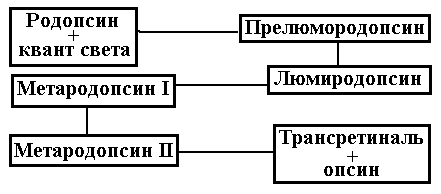
Рис. 6.23. Превращение молекулы
родопсина при поступлении кванта света.
В рецепторных клетках ионы кальция
слабо связаны с метародопсином, либо с
соседними мембранными белками. Запускаемый
цикл фотохимических процессов при
участии кальмодулина активирует ионы
Са2+. Свободный кальций и другие
вторичные посредники (цГМФ) диффундируют
к плазматической мембране, где изменяют
ее проницаемость для Nа+: возникает
рецепторный потенциал (РП) (рис. 6.24). Но
РП, возникающий в фоторецепторах под
действием кванта света, обусловлен не
деполяризацией, как в других рецепторах,
агиперполяризацией мембраны
рецепторной клетки. В темноте мембрана
их сравнительно хорошо пропускает ионы
натрия, что приводит к низкому уровню
ее поляризации (около -25 мВ). При действии
света проницаемость натриевых каналов
снижается, поэтому потенциал мембраны
возрастает. Причем, амплитуда
гиперполяризации растет с увеличением
освещенности (максимально до -90 мВ).
Р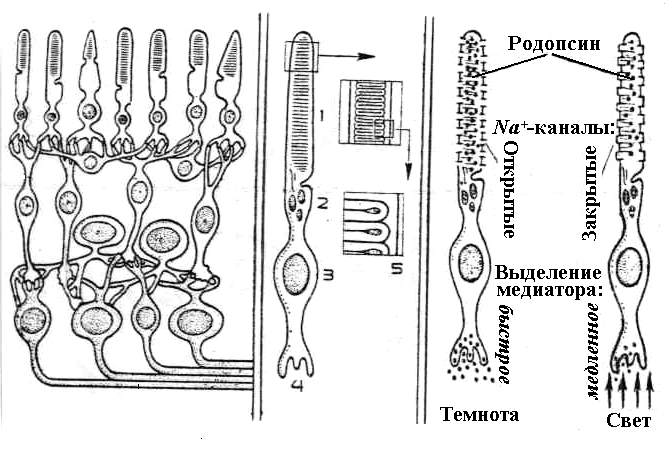 одопсин
одопсин
палочек наиболее чувствителен при
действии волны 505 нм. Цветочувствительные
пигменты колбочек, состоящие из ретиналя
и фотопсина (белковая часть его несколько
отличается от скотопсина палочек), имеют
пики чувствительности: голубочувствительные
— 445 нм, зелено-чувствительые — 535 нм и
красночувствительные при 570 нм (рис.
6.25).
Р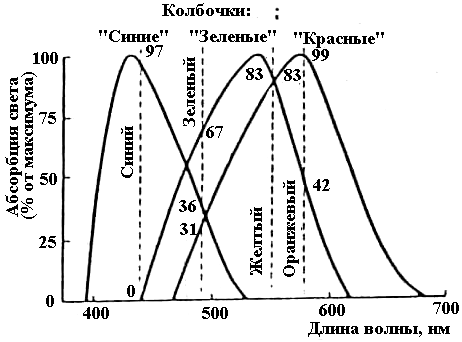 ис.
ис.
6.24.Схема строения сетчатки (слева),
палочка (в центре):1, 5 — мембраны с
фиксированными на них пигментами,2— митохондрии, ядро, 4— синапс.
Рис. 6.25. Спектральная чувствительность
трех типов колбочек (по Mc Nihol).
Рецепторный потенциал в палочках
развивается медленнее, чем в колбочках.
Обусловлено это скорее всего тем, что
в палочках ион кальция должен пройти
большее расстояние до ближайшего
нат-риевого канала, чем в колбочках: в
па-лочках большая часть молекул пигмента
находится в мембране дисков, в то время
как в колбочках — в складках самой
кле-точной мембраны. Поэтому при изменении
освещенности палочковая система более
инерционна (см. «Адаптация рецепторов»).
Обработка зрительной информации в нейронах сетчатки
Нейроны сетчатки включают четыре типа
клеток: горизонтальные, биполярные,
амакриновые и ганглиозные(рис. 6.24).
Фоторецепторные клетки с помощью
синаптических контактов передают
сигналы на биполярные клетки. Биполярные
клетки, в свою очередь, через синапс,
расположенный на другом полюсе их,
передают возбуждение на дендриты
ганглиозных клеток. Горизонтальные
клетки объединяют несколько синапсов
биполярных клеток с фоторецепторами,
а амакриновые клетки — синапсы биполярных
клеток с ганглиозными. Так как биполярные,
горизонтальные и амакриновые нейроны
имеют настолько малые размеры то,
возбуждение онимогут проводить
пассивно в виде РП, без возникновения
потенциала действия.Потенциал
действия возникает лишь на уровне
ганглиозных клеток,дающих начало
зрительному нерву.
В нейронах сетчатки при передаче сигналов
широко происходят процессы конвергенции
и дивергенции. Биполярные клетки
объединяют несколько фоторецепторов,
а каждая ганглиозная клетка на входе
получает импульсы от нескольких
биполярных клеток. В результате происходит
конвергенция (схождение) зрительных
стимулов. В конвергенции ведущее
значение принадлежит горизонтальным
и амакриновым клеткам, которые
ответственны за передачу сигналов
латерального торможения.
Степень дивергенции(расхождения)
зависит от величины дендритного дерева
ганглиозной клетки и контактирующих с
ней нейронов. В центральной ямке и вблизи
нее колбочки и палочки через биполярные
клетки контактируют преимущественно
с индивидуальными ганглиозными клетками.
Это обеспечивает высокую остроту зрения
данного отдела сетчатки. Чем дальше к
периферии сетчатки, тем степень
дивергенции ганглиозных клеток более
выражена. На периферии до 300 палочек
конвергирует на одну ганглиозную клетку,
что обеспечивает суммацию ВПСП. В
результате такойсуммациипри
расширении зрачка чувствительность
глаза в сумерках повышается. Хотя при
этом одновременно снижается острота
зрения. Суммарно в сетчатке преобладают
процессы конвергенции над дивергенцей,
о чем свидетельствует несоответствие
рецепторных клеток (125 млн.) афферентным
нейронам гангиозных клеток (1 млн.).
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Рецепторные клетки сетчатки.Сетчатка
является внутренней оболочкой глаза
(рис. 6.21). Здесь располагаются фоторецепторы
(палочки и колбочки), несколько видов
нервных клеток и слой пигментных клеток.
Светочувствительные членики фоторецепторов
обращены в сторону пигментных клеток.
В центре сетчатки можно выделить два
образования: 0,4 мм диаметромцентральная
ямка(fovea centralis) ислепое пятно—
место входа зрительного нерва.
Прошедший через оптическую систему
глаза свет попадает на сетчатку, где
воспринимается рецепторными клетками.
Слой рецепторов в глазу человека состоит
приблизительно из 120 млн. палочек и 6
млн. колбочек. Они отличаются даже по
внешнему виду (рис. 6.21), по местоположению
на сетчатке и функциональному назначению.
Максимальная плотность колбочек в
центре fovea.Наибольшая плотность
палочек в парафовеальной области, а в
самой центральной ямке палочки полностью
отсутствуют(рис. 6.22).
Рис. 6.21. Схема расположения клеточных
элементов сетчатки: слева — на периферии,
справа — в центре.
Каждый фоторецептор состоит из
светочувствительного наружного сегмента,
содержащего зрительные пигменты и ядро;
внутреннего сегмента, содержащего
большое количество митохондрий и других
субклеточных структур; синаптического
тела. Наружный сегмент палочки состоит
примерно из 400-800 тонких дисковидных
пластинок. Диск представлен двойной
мембраной, с молекулами белка которой
связан зрительный пигмент — родопсин.
Примерно такую же структуру имеет и
наружный сегмент колбочек, но в них
мембранные структуры образуют складки.
С мембраной складок связаны зрительные
пигменты. Колбочки различаются по
наличию трех типов зрительного пигмента:
иодопсина, хлоролаба и эритролабаСтруктура всех зрительных пигментов
очень близка, но имеющееся небольшое
различие ее обеспечивает специфическую
чувствительность каждого из них к
действию определенной длины волны (см.
ниже).
Диаметр наружного сегмента палочек от
2 до 5 мкм, а колбочек от 5 до 8 мкм.
Пигментный слой. В пигментном слое
сетчатки содержится черный пигмент —меланин, принимающий активное
участие в обеспечении ясного видения.
Пигмент, поглощая свет, препятствует
его отражению от стенок и попаданию на
другие рецепторные клетки. Участие
пигмента наглядно видно у альбиносов,
людей с врожденным отсутствием его. У
них через зрачок видны красные сосуды.
В ярко освещенном помещении альбинос
теряет способность ясно различать
предметы. Кроме того пигментный слой
содержит большое количествовитамина
А, участвующего в ресинтезе зрительных
пигментов в наружных сегментах палочек
и колбочек,куда он может легко
передаваться. Эти процессы особенно
интенсивно совершаются в палочках.
Поэтому при недостаточном поступлении
в организм витамина А может развиться,
так называемая,куриная слепота—
нарушение зрения при плохом освещении,
когда за счет расширения зрачка
присоединяется палочковый аппарат.
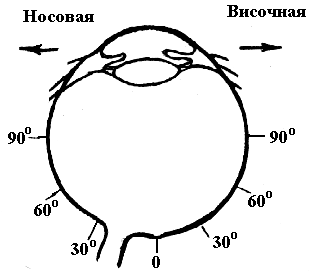
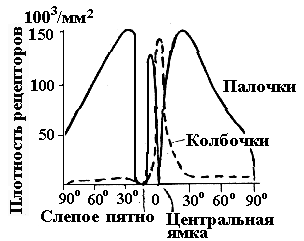
Рис. 6.22.Плотность размещения палочек
и колбочек в различных отделах сетчатки.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Сетчатка это часть мозга, выдвинутая на периферию. Мы рассмотрели
некоторые ее детали от уровня молекул до уровня «блок-схем». Ее функции
прослеживаются вплоть до молекулярного уровня. Хотя остается еще немало
неясного в том, что касается синаптологии и клеточных взаимодействий, мы
явно идем к формированию всесторонней теории — модели сетчатки. Если эти
исследования будут доведены до конца, это будет первой частью мозга,
изучение которой доведено до полного понимания. Значение такой
окончательной теории огромно. Остающиеся проблемы, однако, огромны. Мы
рассмотрели организацию сетчатки по вертикали и горизонтали. Диффузные
биполярные клетки могут вступать в контакт и объединять активность до
десяти ножек колбочек и передавать ее на единственную ганглиозную клетку; с
другой стороны, карликовые биполяры часто образуют с ганглиозными клетками
связи «один к одному», и так передают информацию в волокна зрительного
нерва. Горизонтальные и амакриновые клетки вмешиваются в этот вертикальный
поток информации и модулируют ее передачу. Таким образом, в сетчатке
сочетаются параллельная и иерархическая обработка информации — здесь, на
периферии, уже начинается то, что происходит затем в мозге. Мы увидим далее
в главе ЗРИТЕЛЬНЫЕ ПУТИ , что более двух дюжин
экстрастриарных участков коры приматов соединены «вертикально» и
«перекрестно». В сетчатке мы в миниатюре можем изучать сложную смесь
параллельной и иерархической обработки информации, которая в много более
сложном виде наблюдается в мозге. Но сложности на этом не заканчиваются. В
отличие от однозначных переключателей в устройстве компьютера,
биологические «переключатели» в мозге — это сложный коктейль медиаторов , это ГАМК, глютатмат,
ацетилхолин, дофамин, серотонин, соматостатин и многое другое,
присутствующее здесь одновременно. Кто-то возбуждает, кто-то — тормозит, а
кто-то — модулирует. Рецепторов, с которыми все они взаимодействуют,
большое множество и они различны. Методы современной молекулярной биологии
были применены для клонирования многих из этих крупных рецепторных белков,
и было изучено не только их структурное разнообразие, но и динамика их
синтеза и деградации, которые постоянно меняют восприимчивость
постсинаптических мембран к многокомпонентному коктейлю нейрохимических
веществ. Следует также помнить, что сетчатка человека — это лишь один
случай из многих. Человек — один из приматов, которые тоже разнообразны, а
сами приматы — всего лишь один отряд из числа млекопитающих ( гл. КЛАССИФИКАЦИЯ ОРГАНИЗМОВ ). Ну и сетчатка
млекопитающих — это лишь часть спектра сетчаток позвоночных. Некоторых ее
вариантов мы коснемся в гл. ЗРИТЕЛЬНЫЕ СИСТЕМЫ
ДРУГИХ ПОЗВОНОЧНЫХ и увидим, что на основе сравнительно
неспециализированной сетчатки, подобной нашей, развились удивительные
модификации у глубоководных рыб и хищных птиц.
Есть однако черта, по
которой сетчатка приматов превосходит остальных млекопитающих (хотя и не
всех позвоночных) — это цветное зрение . Мы
видели в разделе Фоторецепторные клетки , что
цветное зрение в сетчатке человека зависит (исходно) от наличия трех
различных пигментов колбочек. Приматы — единственные среди млекопитающих, у
кого пигментов — три, у остальных — только два. Грубо говоря, у этих
животных один пигмент — коротковолновый, а другой — длинноволновый. Собака
— типичный дихромат, один из пигментов колбочек имеет max = 450 нм, а
другой — max = 555 нм. Домашняя кошка — тоже дихромат с коротко- и
длинноволновым пигментами, у нее, однако, пока не удалось выявить цветное
зрение поведенческими экспериментами. Почему же люди и другие приматы
оказались единственными трихроматами среди млекопитающих? Наиболее
правдоподобное объяснение состоит в том, что трихромазия развилась у
существ, обитающих под пологом леса, под давлением необходимости правильно
оценивать спелость (или, соответственно, незрелость) тропических фруктов.
Как мы видели в разделе Горизонтальные клетки ,
то эволюционное событие, которое приведшело к возникновению опсина зеленых
колбочек, занявшего в спектре промежуточное положение между длинноволновым
(«красным») и коротковолновым («синим») опсинами, произошло около 40
миллионов лет назад. Тогда ранние приматы адаптировались к преимущественно
древесному образу жизни и фруктовой диете. Когда эта книга уже была
направлена в печать, появились сообщения о трихомазии у дневных prosimian
(скользяще-лазающие лемуры, медленный лори) — далеких родичей человека,
высших и низших обезьян. Хотя сетчатки других млекопитающих не могут
конкурировать с сетчаткой приматов в количестве колбочковых пигментов, они
часто специализировались иными путями. Во многих случаях, как у кролика и
тем более у многих позвоночных, не относящихся к млекопитающим, обработка
информации в большей степени, чем у приматов, осуществляется в сетчатке. В
сетчатке кролика есть ганглиозные клетки, способные реагировать на большое
разнообразие стимулов, а не просто на пятно света, как сетчатка приматов
или кошек. Обнаружены ганглиозные клетки, активируемые движущимися краями
объектов (локальные детекторы краев), краев специфической ориентации,
движущихся в определенных направлениях, а также клетки, чьи рецептивные
поля настроены на выделение других, часто очень специфических, ключевых
стимулов. У приматов и кошек клетки, отвечающие на такие специфические
стимулы, находятся в зрительной коре, и мы обсудим некоторые из таких
случаев в следующей главе. Возможно, дело в том, что кролику необходимо
экстренно обрабатывать ту информацию, которую другие животные, на которых
охотятся не так часто, могут обдумать не спеша. Именно это и происходит в
анализаторе зрительной коры приматов и кошачьих, к которым мы перейдем в
следующей главе. Фоторецепторы сетчатки:
обновление
Смотрите также:
Источник
