
Горизонтальная клетка в сетчатке
Функция горизонтальных клеток сетчатки. Возбуждение и торможение биполярных клетокГоризонтальные клетки, показанные на рисунке, обеспечивают связь между синаптическими телами палочек и колбочек и дендритами биполярных клеток. На выходе горизонтальные клетки всегда тормозные. Следовательно, эта латеральная связь обеспечивает феномен латерального торможения, характерный для всех других сенсорных систем и важный для повышения надежности проведения визуального образа с соответствующим зрительным контрастом. Этот феномен демонстрируется на рисунке, где показано небольшое пятно света, сфокусированное на сетчатке. Зрительный путь от самой центральной области, куда попадает световой луч, возбуждается, тогда как рядом лежащая область тормозится. Другими словами, горизонтальные клетки путем латерального торможения в окружающих областях останавливают диффузное распространение сигнала по сетчатке, которое могло бы возникнуть в связи с наличием широкого ветвления дендритов и аксонов в слоях сетчатки. Это важно для четкого выделения контрастных границ в зрительном образе. Возможно, что некоторые амакриновые клетки также обеспечивают дополнительное латеральное торможение и дальнейшее усиление зрительного контраста во внутреннем слое сетчатки. Возбуждение и торможение биполярных клетокПротивоположные сигналы (возбуждающие и тормозные) в зрительном пути обеспечиваются двумя типами клеток: (1) деполяризующимися биполярными клетками (2) гиперполяризующимися биполярными клетками. Это значит, что при возбуждении палочек и колбочек некоторые биполярные клетки деполяризуются, тогда как другие гиперполяризуются. Существуют два возможных объяснения этого различия. Согласно одному из них имеются биполярные клетки двух типов, которые по-разному реагируют на выделяемый палочками и колбочками медиатор глутамат: одни деполяризуются, другие гиперполяризуются. Другим возможным объяснением является то, что одни биполярные клетки получают сигнал непосредственно от палочек и колбочек, тогда как другие получают этот же сигнал через горизонтальные клетки. Поскольку горизонтальные клетки тормозные, их действие приводит к противоположной реакции биполярных клеток. Независимо от механизма разнонаправленных реакций, важность этого феномена состоит в том, что одна половина биполярных клеток проводит положительные сигналы, а другая — отрицательные. Как мы увидим далее, оба типа сигналов — и положительный, и отрицательный — используются при проведении зрительной информации в мозг. Другим важным аспектом этого реципрокного отношения между деполяризующимися и гиперполяризующимися биполярными клетками является обеспечение второго механизма латерального торможения дополнительно к механизму, связанному с горизонтальными клетками. Поскольку разные типы биполярных клеток лежат рядом друг с другом, это способствует появлению разделяющих контрастных границ в зрительном образе, даже если граница лежит точно между двумя прилежащими фоторецепторами. В противоположность этому механизм латерального торможения, обеспечиваемый горизонтальными клетками, действует на гораздо большем расстоянии.
— Также рекомендуем «Амакриновые клетки. Ганглиозные клетки сетчатки» Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»: |
Источник
Горизонтальные клетки сетчатки глаза — H — располагаются сразу за фоторецепторами R,C и отдают им большое количество дендритов, которые, переплетаясь, образуют сплошное густое кружево. Их аксоны могут тянуться на значительные расстояния и имеют сильно разветвленные концевые образования. Горизонтальные клетки соединены между собой электрическими синапсами и образуют функциональный синцитий. Возбуждение одной горизонтальной клетки распространяется на другие. В центральной ямке (лат. fovea) сетчатки приматов одна горизонтальная клетка соединяется примерно с 6-ю колбочками, а на периферии — с 30-40.[2]Дендритное поле горизонтальной клетки на периферии сетчатки достигает диаметра 250 мкм. В то же время, размер рецептивных полей горизонтальных клеток может изменяться в пределах 300мкм ÷ 5мм[3], что значительно превышает протяженность дендритных разветвлений. За счет этого горизонтальные клетки могут формировать периферию рецептивных полей биполярных нейронов[4][5][6][7][8][9][10]. При освещении фоторецепторов горизонтальные клетки отвечают гиперполяризациейГиперполяризация — увеличение разности потенциалов между наружной и внутренней сторонами биологической мембраны в возбудимых тканях.. (Увеличение разности потенциалов между наружной и внутренней сторонами биологической мембраны в возбудимых тканях).

Рис.14. Наслоение и выключение биполярных аксонов клеток кошки внутреннего плексиформного слоя (IPL). OFF — (отключение) ганглиозных клеток ( αGC и βGC ) дендритных и конусных мембран колбочек OFF биполярных аксонов клеток ( OFF CB ) совместно слоиться в sublamina из IPL. НА (включение) — биполярных аксонов (O N CB ) и ганглиозных клеток дендритов со-стратификацией в sublamina б в IPL. Это параллельные включения и выключения колбочковых путей, которые происходят с биполярными клетками дендритных контактов с мембранами колбочекко. На рисунке изменяется от Nelson и соавт, 1978 (100).[2]
Учитывая связь горизонтальных клеток H с колбочками (cones) (см. рис.14) в центральной ямке сетчатки глаза, где нет палочек, они активно участвуют в цветном зрении.
Фоторецепторы вместе с горизонтальными клетками и биполярами образуют так называемую триаду[6]. Триада состоит из пресинаптической мембраныПресинаптическая мембрана — утолщенный участок мембраны синаптической бляшки в области синапса. (утолщенный участок мембраны синаптической бляшки в области синапса) фоторецептора, в которую заходит дендрит биполяра, а по бокам от него заходят отростки горизонтальных клеток. Значение и принципы функционирования триады пока остаются неизвестными. За счет таких связей между фоторецепторами, горизонтальными клетками и биполярами, центр рецептивного поля биполяров формируется прямым путем при передаче сигнала от фоторецепторов, периферия формируется за счет непрямого пути через горизонтальные клетки за счёт возбуждающих или тормозных синапсов.
См. также[править]
- Цветное зрение
Примечания[править]
- ↑ https://ru.wikipedia.org/wiki/%D0%A4%D0%B0%D0%B9%D0%BB:Retina_layers.svg
- ↑ а б Ralph Nelson and Victoria Connaughton. «bipolar-cell-pathways-in-the-vertebrate-retina». Webvision. Retrieved Лаборатория доктора Марка,28 сентября, 2012.
- ↑ Измайлов И. А., Соколов Е. Н., Чернорызов А. М. Психофизиология цветового зрения. — М.: Изд-во Московского университета, 1989. — 206 с.
- ↑ Школьник-Яррос Е. Г. , Калинина А. В. Нейроны сетчатки. — М.: Наука, 1986. — 208 с.
- ↑ Масланд Р. Функциональная организация сетчатки// В мире науки. — 1987, № 4 — с. 58-68.
- ↑ а б Бызов А. Л. Полищук Н. А. О механизме обратной связи от горизонтальных клеток к фоторецепторам: химеческая или електрическая гипотеза?// Сенсорные системы. — 1987. Т1, № 4. — с.344-352.
- ↑ Бызов А. Л., Голубцов К. В. Модель нейрона-регулятора эффективности синаптической передачи// Биофизика. — 1978. — т.23 № 1. — с. 119—125.
- ↑ Шмидт Р. Тевс Г. Физиология человека. т.1. -М.: Мир, 1996. — 328 с.
- ↑ Dacey D. Lee B. Stafford D. Horizontal cells of the primate retina: cone specifity without spectral opponency // Science. — 1996. Vol. 271, № 5429. р. 656—659.
- ↑ Глезер В. Д. Зрение и мышление. — Л.: Наука, 1985, — 248 с.
Источник
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 8 сентября 2018;
проверки требуют 3 правки.
Запрос «Ретина» перенаправляет сюда; о названии особого вида ЖК-дисплеев см. Retina.
Сетча́тка (лат. retína) — внутренняя оболочка глаза, являющаяся периферическим отделом зрительного анализатора; содержит фоторецепторные клетки, обеспечивающие восприятие и преобразование электромагнитного излучения видимой части спектра в нервные импульсы, а также обеспечивает их первичную обработку.
Строение[править | править код]
Анатомически сетчатка представляет собой тонкую оболочку, прилежащую на всём своём протяжении с внутренней стороны к стекловидному телу, а с наружной — к сосудистой оболочке глазного яблока. В ней выделяют две неодинаковые по размерам части: зрительную часть — наибольшую, простирающуюся до самого ресничного тела, и переднюю — не содержащую фоточувствительных клеток — слепую часть, в которой выделяют в свою очередь ресничную и радужковую части сетчатки, соответственно частям сосудистой оболочки.
Зрительная часть сетчатки имеет неоднородное слоистое строение, доступное для изучения лишь на микроскопическом уровне и состоит из 10[2] следующих вглубь глазного яблока слоёв:
- пигментного,
- фотосенсорного,
- наружной пограничной мембраны,
- наружного зернистого слоя,
- наружного сплетениевидного слоя,
- внутреннего зернистого слоя,
- внутреннего сплетениевидного слоя,
- ганглионарных клеток,
- слоя волокон зрительного нерва,
- внутренней пограничной мембраны.
Строение сетчатки человека[править | править код]
Сетчатка глаза у взрослого человека имеет диаметральный размер 22 мм и покрывает около 72 % площади внутренней поверхности глазного яблока.
Пигментный слой сетчатки (самый наружный) с сосудистой оболочкой глаза связан более тесно, чем с остальной частью сетчатки.
Около центра сетчатки (ближе к носу) на задней её поверхности находится диск зрительного нерва, который иногда из-за отсутствия в этой части фоторецепторов называют «слепое пятно». Он выглядит как возвышающаяся бледная овальной формы зона около 3 мм². Здесь из аксонов ганглионарных нейроцитов сетчатки происходит формирование зрительного нерва. В центральной части диска имеется углубление, через которое проходят сосуды, участвующие в кровоснабжении сетчатки.
диска зрительного нерва, приблизительно в 3 мм, располагается пятно (macula), в центре которого имеется углубление, центральная ямка (fovea), являющееся наиболее чувствительным к свету участком сетчатки и отвечающее за ясное центральное зрение (жёлтое пятно). В этой области сетчатки (fovea) находятся только колбочки. Человек и другие приматы имеют одну центральную ямку в каждом глазу в противоположность некоторым видам птиц, таким как ястребы, у которых их две, а также собакам и кошкам, у которых вместо ямки в центральной части сетчатки обнаруживается полоса, так называемая зрительная полоска. Центральная часть сетчатки представлена ямкой и областью в радиусе 6 мм от неё, далее следует периферическая часть, где по мере движения вперед число палочек и колбочек уменьшается. Заканчивается внутренняя оболочка зубчатым краем, у которого фоточувствительные элементы отсутствуют.
На своём протяжении толщина сетчатки неодинакова и составляет в самой толстой своей части, у края диска зрительного нерва, не более 0,5 мм; минимальная толщина наблюдается в области ямки жёлтого пятна.
Микроскопическое строение[править | править код]
Упрощенная схема расположения нейронов сетчатки. Сетчатка состоит из нескольких слоев нейронов. Свет падает слева и проходит через все слои, достигая фоторецепторов (правый слой). От фоторецепторов сигнал передается биполярным клеткам и горизонтальным клеткам (средний слой, обозначен жёлтым цветом). Затем сигнал передается амакриновым и ганглионарным клеткам (левый слой). Эти нейроны генерируют потенциалы действия, передающиеся по зрительному нерву в мозг. С рисунка Сантьяго Рамон-и-Кахаля, видоизменено
См. Пигментный эпителий сетчатки
В сетчатке имеются три радиально расположенных слоя нервных клеток и два слоя синапсов.
Ганглионарные нейроны залегают в самой глубине сетчатки, в то время как фоточувствительные клетки (палочковые и колбочковые) наиболее удалены от центра, то есть сетчатка глаза является так называемым инвертированным органом. Вследствие такого положения свет, прежде чем упасть на светочувствительные элементы и вызвать физиологический процесс фототрансдукции, должен проникнуть через все слои сетчатки. Однако он не может пройти через пигментный эпителий или хориоидею, которые являются непрозрачными.
Проходящие через расположенные перед фоторецепторами капилляры лейкоциты при взгляде на синий свет могут восприниматься как мелкие светлые движущиеся точки. Данное явление известно как энтопический феномен синего поля (или феномен Ширера).
Кроме фоторецепторных и ганглионарных нейронов, в сетчатке присутствуют и биполярные нервные клетки, которые, располагаясь между первыми и вторыми, осуществляют между ними контакты, а также горизонтальные и амакриновые клетки, осуществляющие горизонтальные связи в сетчатке.
Между слоем ганглионарных клеток и слоем палочек и колбочек находятся два слоя сплетений нервных волокон со множеством синаптических контактов. Это наружный плексиформный (сплетеневидный) слой и внутренний плексиформный слой. В первом осуществляются контакты между палочками и колбочками и вертикально ориентированными биполярными клетками, во втором — сигнал переключается с биполярных на ганглионарные нейроны, а также на амакриновые клетки в вертикальном и горизонтальном направлении.
Таким образом, наружный нуклеарный слой сетчатки содержит тела фотосенсорных клеток, внутренний нуклеарный слой содержит тела биполярных, горизонтальных и амакриновых клеток, а ганглионарный слой содержит ганглионарные клетки, а также небольшое количество перемещённых амакриновых клеток. Все слои сетчатки пронизаны радиальными глиальными клетками Мюллера.
Наружная пограничная мембрана образована из синаптических комплексов, расположенных между фоторецепторным и наружным ганглионарным слоями. Слой нервных волокон образован из аксонов ганглионарных клеток. Внутренняя пограничная мембрана образована из базальных мембран мюллеровских клеток, а также окончаний их отростков. Лишённые шванновских оболочек аксоны ганглионарных клеток, достигая внутренней границы сетчатки, поворачивают под прямым углом и направляются к месту формирования зрительного нерва.
Каждая сетчатка у человека содержит около 6—7 млн колбочек и 110—125 млн палочек. Эти светочувствительные клетки распределены неравномерно. Центральная часть сетчатки содержит больше колбочек, периферическая содержит больше палочек. В центральной части пятна в области ямки колбочки имеют минимальные размеры и мозаично упорядочены в виде компактных шестиграных структур.
Заболевания[править | править код]
Есть множество наследственных и приобретённых заболеваний и расстройств, поражающих, в том числе, сетчатку. Перечислены некоторые из них:
- Пигментная дегенерация сетчатки — наследственное заболевание с поражением сетчатки, протекает с утратой периферического зрения.
- Дистрофия жёлтого пятна — группа заболеваний, характеризующихся утратой центрального зрения вследствие гибели или повреждения клеток пятна.
- Дистрофия макулярной области сетчатки — наследственное заболевание с двусторонним симметричным поражением макулярной зоны, протекающее с утратой центрального зрения.
- Палочко-колбочковая дистрофия — группа заболеваний, при которых потеря зрения обусловлена повреждением фоторецепторных клеток сетчатки.
- Отслоение сетчатки от задней стенки глазного яблока. Игнипунктура — устаревший метод лечения.
- И артериальная гипертензия, и сахарный диабет могут вызвать повреждение капилляров, снабжающих сетчатку кровью, что ведёт к развитию гипертонической или диабетической ретинопатии.
- Ретинобластома — злокачественная опухоль сетчатки.
- Меланома сетчатки- злокачественная опухоль из пигментных клеток- меланоцитов, рассеянных в сетчатке.
- Макулодистрофия — патология сосудов и нарушение питания центральной зоны сетчатки.
Литература[править | править код]
- Савельева-Новосёлова Н. А., Савельев А. В. Принципы офтальмонейрокибернетики // В сборнике «Искусственный интеллект. Интеллектуальные системы». — Донецк-Таганрог-Минск, 2009. — С. 117—120.
Примечание[править | править код]
Ссылки[править | править код]
- Строение сетчатки. // Проект «Eyes for me».
Источник
Описание

Электронно-микроскопические наблюдения последних лет, а также электрофизиологические микроэлектродные исследования позволили окончательно установить, что горизонтальные клетки — нейроны и отвергнуть гипотезу о глиальной их природе [Максимова, 1970; Бызов, 1971, 1984; Gouras, 1972; Rodieck, 1973; Gallego, 1976; Naka, 1982].
Сходная дискуссия происходила еще в прошлом веке, когда ряд ученых считали горизонтальные клетки опорными или поддерживающими, a Ramon у Cajal [1972] не сомневался, что они типичные нейроны.
Роль горизонтальных клеток полностью не ясна, но совершенно очевидно, что они мультифункциональны. Топографическое положение синаптических окончаний отростков этих нейронов в триадах фоторецепторов обусловливает возможность влияния их на синаптическую передачу от рецептора к биполяру [Бызов, Голубцов, 1978; Byzov, 1979; Бызов, 1984]. Как интернейроны в других отделах нервной системы, горизонтальные клетки воспринимают сигналы от одних элементов — фоторецепторов и передают их различным структурам сетчатки — другим горизонтальным клеткам, обратно фоторецепторам, биполярным клеткам, опосредованно — ганглиозным клеткам.
Соотношение инвагинирующей в фоторецептор биполярной клетки с горизонтальными особенно ясно и детально представлено Sjostrand [1974, 1976]. По данным Sjostrand, от основного инвагинирующего отросточка биполяра отходит крошечное ответвление, входящее в «шейку» синаптического ленточного комплекса фоторецептора и являющееся постсинаптическим к отросткам горизонтальных клеток (см. рис. 3).
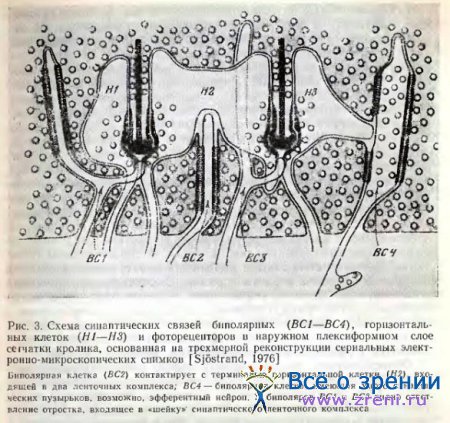
Автор считает, что процесс адаптации зависит от тормозного действия горизонтальных клеток на биполярную, осуществляющегося в «шейке» синаптического комплекса фоторецептора.
По Barlow, Levick [1965], горизонтальные клетки играют роль в организации деятельности детекторов направления движения. Из ряда физиологических работ следует, что горизонтальные клетки организуют периферию рецептивных потей ганглиозных клеток [Максимова, 1969; Naka, Nye, 1971; и др.]. Тела горизонтальных клеток ограничивают с внутренней (витреальной) стороны наружный плексиформный слой; отростки пронизывают его, направляясь к фоторецепторам. Продемонстрированы реакции горизонтальных клеток разных слоев: реакции L-тииа генерируются в ближнем к рецепторам слое, хроматические реакции R/G-типа — в более удаленном слое. Mitarai, Asa-no, Miyake [1974] подтвердили многообразие типов горизонтальных нейронов в сетчатке рыб. Авторы идентифицировали 5 типов потенциалов, возникающих в различных слоях клеток. Фотопические реакции L-типа генерируются в наружных горизонтальных клетках. Потенциалы С-типа — красно-зеленые, трехфазные и желто-синие — возникают в средних горизонтальных клетках. Идентификация нейронов проводилась при помощи внутриклеточных меток. Свойства рецептивных полей горизонтальных клеток, возможно, зависят от активности интерплексиформных клеток. Так, разрушение дофаминергических терминалей, свойственных интерплексиформным клеткам и оканчивающихся в наружном плексиформном слое, приводило к увеличению рецептивного поля горизонтальных клеток у карпа [Cohen, Dowling, 1983].
Морфология горизонтальных клеток весьма разнообразна в зависимости от вида исследуемого представителя позвоночных. У рыб наиболее крупные нейроны, образующие подслои или ряды.


У щуки (см. рис. 1, А) три ряда занимают почти всю ширину внутреннего ядерного слоя. Parthe [1972] в превосходной работе весьма убедительно демонстрирует у Mugil brasiliensis, Eugerres plumieri и других рыб четыре типа горизонтальных клеток — три типа колбочковых и один тип палочковых. У Саrassius auratus, как следует из весьма детального исследования Stell [1975], Stell, Lightfoot [1975], несмотря па наличие лишь узкой прослойки сетчатки в 5—10 мкм толщиной, занятой горизонтальными нейронами, они также четко дифференцированы на три колбочковых и один палочковый типы. Различия в типах касаются размеров тел, обширности ветвлений дендритов и характера синаптических связей. Три типа колбочковых горизонтальных клеток у Carassius auratus обозначены Stell HI, H2, Н3. Каждому типу соответствует особая морфология, сочетающаяся со свойственной лишь этому типу функциональной характеристикой.
Столько же типов горизонтальных клеток в сетчатке черноморской ставриды [Подугольникова, Максимов, 1984]; в сетчатках очень большого количества разнообразных видов костистых рыб три их типа и соответственно ряда [Wagner, 1972; Подугольникова, 1981]. Но у некоторых рыб всего два ряда или даже лишь один ряд горизонтальных клеток, например у зеленушки [Подуголышкова, 1981]. Горизонтальные клетки в сетчатке рыб образуют правильные решетки, элементы которой расположены регулярно, с определенной периодичностью по отношению к периодичности расположения фоторецепторов [Wagner, 1972; Подуголышкова, Максимов, 1973, 1977]. Горизонтальные клетки одного и того же ряда соединяются друг с другом посредством тесных контактов [Yamada, Ishikawa, 19G5; Боровягин, 1966].
Применив Гольджи — электронную микроскопию, Stell, Lightfoot [1975] выяснили избирательность связей определенного типа горизонтальных клеток с колбочками. Так, клетки III-типа, отличающиеся наибольшей величиной тела и наименьшим размахом дендритов, функционально связаны с красно-чувствительными колбочками. Клетки Н2-типа никогда не контактируют с длинными членами двойных колбочек и с длинными одиночными колбочками. Функционально они связаны с зелено- и синечувствительными колбочками. Н3-клетки, имеющие небольшое тело и широкий размах дендритов, контактируют лишь с короткими и миниатюрными одиночными колбочками, т. с. с синечувствительными.
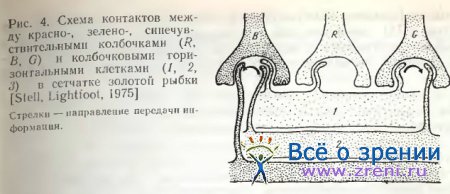
Stell [1975] также указывает на возможные пути передачи сигналов (рис. 4). Так, через Н1-клетки красночувствительные колбочки связываются с Н2-клетками, через Н2-клетки зелено-чувствительные колбочки и Н1-клетки связываются с Н3-клетками. В сетчатке ставриды в отличие от сетчатки золотой рыбки горизонтальные клетки среднего слоя контактируют только с красночувствительными двойными колбочками [Подугольникова, 1981]. У всех колбочковых горизонтальных клеток Carassius auratus Stell [1975] нашел аксоны, спускающиеся в глубь внутреннего ядерного слоя, где они, простираясь в горизонтальном направлении, оканчиваются длинными утолщениями. Эти терминальные утолщения аксона настолько велики, что Ramon у Cajal [1972] принимал их за самостоятельный ряд клеток. У палочковых горизонтальных клеток рыб аксоны не обнаружены.
Терминали аксонов горизонтальных клеток, а также связи по ходу аксонов у многих других видов рыб тщательно прослежены методом Гольджи Parthe [1982]. У наружных горизонтальных клеток (Н1) аксон тонкий, варикозный, затем утолщается и с изгибами спускается и переходит в тонкий отросток с отчетливой пуговкой на конце, примыкающей к телу малой биполярной клетки.
По ходу аксона прослеживаются тонкие нитеобразные отросточки с пуговками, оканчивающимися также вблизи тел биполярных клеток. Аксон горизонтальной клетки Н2 у карповых рыб отходит от одного из больших дендритов, в остальном сходен с аксоном у H1. У внутренних горизонтальных клеток Н3 Parthe видел, помимо основных контактов с фоторецепторами, тонкие отросточки дендритов, оканчивающиеся вблизи тел биполярных клеток. У палочковых горизонтальных клеток импрегнировались два типа отростков — многочисленные толстые восходящие, многократно делящиеся и оканчивающиеся у палочковых сферул; глубокие топкие отростки прослеживаются в горизонтальном направлении и оканчиваются крошечными пуговками вблизи тел крупных биполярных клеток.
Горизонтальные клетки сетчатки лягушки весьма полно им-прегнированы по Гольджи еще в прошлом веке Ramon у Cajal [1972]. У малых наружных горизонтальных клеток тонкие длинные дендриты и один наиболее топкий очень далеко прослеживающийся отросток, короткие веточки которого оканчиваются небольшими пуговками у ножек фоторецепторов. Терминали этого аксона — 2-3 варикозные восходящие веточки. Внутренние горизонтальные клетки большей величины с короткими многочисленными дендритами, оканчивающимися у ножек фоторецепторов небольшими варикозными ветвлениями. Отчетливый аксон у внутренних горизонтальных клеток также был обнаружен Ramon у Cajal.
Современные исследователи Hassin, Witkovsky [1983], применив метод Гольджи и введение внутриклеточно люциферового желтого у шпорцевой лягушки (Xenopus), полностью подтвердили классические наблюдения о двух типах нейронов и наличии у обоих типов аксонов. По измерениям этих авторов, у малых клеток диаметр тела — 15-17 мкм, дендритные поля — до 50 мкм. У больших клеток соответственно — 20—25 мкм и до 65 мкм. Аксон прослеживался до 800 мкм без густого терминального ветвления. Электрофизиологически определено, что константа длины горизонтальных клеток сетчатки Xenopus превосходит во много раз величину дендритного ветвления (до 15 раз). Функционально горизонтальные клетки связаны с колбочками и палочками (но не с зелеными палочками).
Таким образом, архитектоника (см. рис. 1) и структурная организация горизонтальных клеток в сетчатке многих рыб и некоторых амфибий весьма различна. Можно думать, что структура слоя горизонтальных; клеток этих животных находится в определенной связи с приспособленностью их зрительного анализатора к среде обитания.
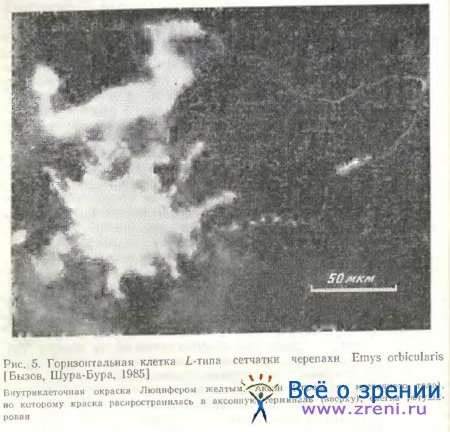
У некоторых видов черепах также обнаруживается 4 типа горизонтальных клеток, но лишь у одного из них (L-типа) найден аксон [Leeper, 1978а]. На рис. 5 представлена горизонтальная клетка сетчатки черепахи Emys orbicularis, окрашенная с помощью внутриклеточного введения люциферового желтого А. Л. Бызовым. Клетка расположена в наружной зоне внутреннего ядерного слоя и относится к типу L. От тела прослеживается тонкий извилистый аксон (справа) и утолщенная терминаль (вверху слева).
Ramon у Cajal различал у всех млекопитающих два типа горизонтальных клеток: наружные, плоские с дендритами, распространяющимися по наружному плексиформному слою, и внутренние, значительно больших размеров. Внутренние горизонтальные клетки подразделяются им далее на имеющие и не имеющие крупные дендритические отростки.
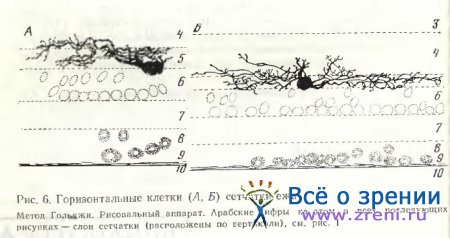
В сетчатке ежа, по нашим предварительным данным,— две разновидности горизонтальных клеток (рис. 6). У более крупных клеток тело плавно переходит в очень широкие густоветвящиеся дендриты, покрытые отчетливыми варикозностями. По ходу тонких отросточков, отходящих от основных дендритов, и на их концах прослеживаются нежные синаптические образования. У горизонтальных клеток меньшей величины тело округлой формы, отростки тоньше и более равномерны по диаметру, чем у крупных клеток. Шипообразные отросточки короче и многочисленнее с типичными синаптическими пуговками на концах. По форме и расположению клеточных тел выявленные нейроны в сетчатке ежа сходны с наружными горизонтальными клетками у млекопитающих по классификации Ramon у Cajal [1972]. В одном случае удалось проследить длинный аксон горизонтальной клетки, распространяющийся по наружному плексиформному слою (рис. 7).

Несколько аксонов у одного из двух типов горизонтальных клеток обнаружил методом Гольджи в сетчатке тупайи (Tupaia glis) Mariani [1985]. Впервые в сетчатке позвоночных автор увидел от 4 до 8 тонких аксонов, начинающихся от концов дендритов. У второго типа горизонтальной клетки морфология отростков не отличается от обычной.
В сетчатке кролика также наблюдаются более крупные и более мелкие горизонтальные клетки. От дендритов горизонтальных клеток в сетчатке ежа и кролика синаптические образования отходят не только по направлению к наружному плексиформному слою, но также в направлении внутреннего ядерного слоя (см. рис. 6). В этих случаях есть основание предполагать существование синаптических связей с отростками других горизонтальных клеток и с отростками биполярных нейронов. Наконец, нельзя исключить возможность связей с телами как горизонтальных, так и биполярных клеток. Действительно электронно-микроскопически синапсы обнаружены между отростками горизонтальных клеток и отростками и телами биполярных клеток в сетчатке кролика и кошки [Dowling, Brown, Major, 1966; Fisher, Boycott, 1974].
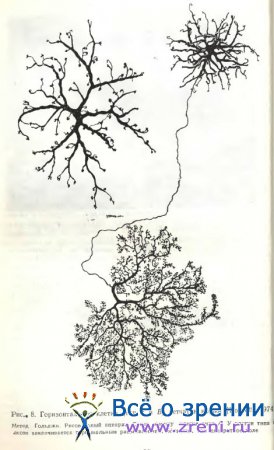
В сетчатке кошки методом Гольджи выделено два типа горизонтальных клеток (рис. 8). Один тип характеризуется отсутствием аксона; у другого аксон обнаружен [Gallego, 1971а; Fisher, Boycott, 1974; Kolb, 1974]. Наиболее детально морфология этих двух разновидностей представлена Fisher, Boycott [1974]. У горизонтальной клетки без аксона дендритное поле достигает на периферии сетчатки 250 мкм в диаметре, дендриты с отходящими от них топкими отросточками оканчиваются скоплениями терминалей. Авторы обозначили подобный нейрон А — типом. У клетки В — типа дендритное поле меньших размеров, отростки тоньше; аксон отходит от одного из дендритов. Терминальное ветвление аксона очень обширное и весьма густое, состоящее из бесчисленного множества мельчайших синаптических пуговок. Подобное терминальное ветвление аксона горизонтальной клетки ранее было обнаружено в сетчатке быка [Ramon у Cajal, 1972], теленка [Marenghi, 1900], собаки [Ramon у Cajal, 1972; Школьник-Яррос, 1958].
Связи дендритов горизонтальных клеток А- и В — типов с фоторецепторами в сетчатке кошки оказались одинаковыми: их окончания образуют латеральные компоненты триад в колбочковых синапсах [Kolb, 1974]. Аксонные окончания у В — типа образуют латеральные компоненты триад в палочковых синапсах. Таким образом, одна горизонтальная клетка В — типа функционально как бы разобщена на палочковую и колбочковую части. Kolb высказывает предположение о возможном смешении палочковых и колбочковых сигналов посредством щелевых контактов, обнаруженных Raviola, Gilula [1973] и другими авторами на уровне самих фоторецепторов, т. е. до их синапсов с горизонтальными клетками. Некоторые различия между двумя типами горизонтальных клеток обнаружены и на ультраструктурном уровне [Fisher, Boycott, 1974]. У клеток A-типа дендриты содержат нейрофиламенты, у клеток В — типа их нет.
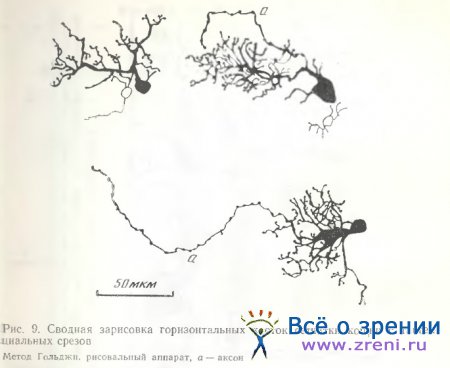
На тангенциальных срезах сетчатки кошки нам удалось также наблюдать аксоны у некоторых горизонтальных клеток (рис. 9). Они отходят от одного из дендритов, как и в случаях, описанных Fisher, Boycott [1974]. По ходу аксонов наблюдаются типичные для аксонов в центральной нервной системе варикозности. У одной из импрегнированных клеток (рис. 9) аксон не виден. Однако известно, что метод Гольджи не дает возможности с полной уверенностью судить об отсутствии какого-либо нейрона или его части, в данном случае аксона. Терминальные ветвления аксонов у импрегнированных нами клеток не выявились. У клетки без аксона дендриты толще; возможно, что это A-тип по Fisher, Boycott [1974]. У всех зарисованных клеток заметно асимметричное расположение дендритов. Асимметрия в расположении дендритов горизонтальных клеток может соответствовать асимметрии рецептивных полей. По мнению Normann, Kolb [1980], определенная ориентация дендритных ветвлений горизонтальных клеток и их рецептивных полей может представлять начальные этапы формирования чувствительности к ориентации, полностью развитой в проксимальных отделах сетчатки и высших зрительных центрах.

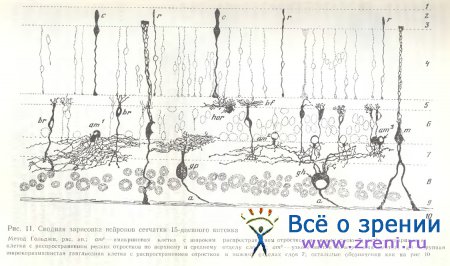
На радиальных срезах сетчатки кошки наблюдаются тела горизонтальных клеток различной формы (рис. 10) и разного размаха ветвления дендритов. Дендритные веточки оканчиваются множеством синаптических пуговок, в наружном плексиформном слое часть из них явно направляется к фоторецепторам, другие же находятся на уровне распространения дендритов биполярных клеток, а иногда и тел других горизонтальных и биполярных клеток. У 15-дневного котенка заметно незначительное еще распространение дендритов горизонтальной клетки (рис. 11) по сравнению со значительным их размахом у 2-месячного котенка.

В сетчатке собаки удалось импрегнировать большое удлиненное веретенообразной формы тело горизонтальной клетки (рис. 12) с обилием дендритов довольно крупных, отходящих от тела почти во все стороны, снабженных большим количеством шиповидных отростков с крупными пуговчатыми синаптическими окончаниями. Следует заметить, что сходство представленной нами клетки из сетчатки собаки с внутренней горизонтальной клеткой сетчатки быка [Ramon у Cajal, 1972; рис. 13, табл. VI] очень велико. Аксоны горизонтальных клеток в сетчатке собаки на своем пути к фоторецепторам пересекают соседние аксоны, а конечные их образования, типичные телодендрин, переплетаясь, образуют сплошное, густое и затейливое кружево (рис. 13).
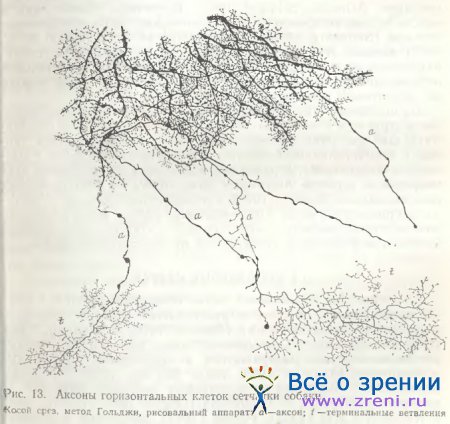
Мелкие синаптические пуговки расположены между крупными стволами аксона.
Ячейки, образующиеся между основными ветвями, часто по форме соответствуют телам горизонтальных клеток (рис. 13) [Школьник-Яррос, 1958]. Образованное телодендриями густое кружево может способствовать взаимным связям между аксонами различных нейронов. При электронно-микроскопическом исследовании аксонных терминалей горизонтальных клеток сетчатки птиц действительно выявляется сеть с многочисленными щелевыми контактами. В этих щелевых контактах, однако, постоянно видны и синаптические пузырьки с обеих сторон [Gallego, 1982]. Типичные щелевые контакты наблюдались также между аксонами горизонтальных клеток в сетчатке шпорцевой лягушки [Witkovsky, Powell, 1981]. Проблема — передаются ли пот

