
Зрительный путь на сетчатке глаза
а) Зрительный нерв, зрительный путь. Зрительный нерв образован аксонами ганглиозных клеток сетчатки. Аксоны покрыты миелиновой оболочкой в месте их выхода из диска зрительного нерва.
Количество ганглиозных клеток значительно варьирует у разных людей и составляет в среднем 1 млн. Поскольку каждая ганглиозная клетка входит в зрительный нерв, количество аксонов в нем соответственно изменяется.
Ганглиозные клетки сетчатки имеют общее происхождение с чувствительными нейронами спинного мозга. Зрительный нерв гомологичен белому веществу спинного мозга и не является периферическим нервом. В главе 9 отмечено, что истинные периферические, черепные или спинномозговые нервы содержат шванновские клетки, покрыты коллагеновой оболочкой и способны к регенерации. Зрительный нерв содержит клетки нейроглии центрального типа (астроциты и олигодендроциты) и не способен к регенерации у млекопитающих. Кроме того, зрительный нерв покрыт мозговыми оболочками с расположенным между ними подпаутинным пространством. Подобное строение объясняет изменения на поверхности глазного дна при повышении внутричерепного давления (отек диска зрительного нерва).
В зрительном перекресте волокна от назальной (медиальной) половины сетчатки входят в противоположный зрительный путь, а волокна от височной (латеральной) половины сетчатки не переходят на другую сторону и попадают в зрительный путь той же стороны. Информация от сетчатки поступает к среднему мозгу (участвующему в регуляции движений глаз, размера зрачка и циркадных ритмов) и латеральному коленчатому ядру таламуса (откуда направляется к зрительной коре, отвечающей за различные аспекты зрения) от различных групп ганглиозных клеток.
Часть зрительных волокон входит в надперекрестное ядро гипоталамуса («центральные часы»), отвечающее за поддержание циркадных ритмов. Такая связь позволяет объяснить положительный эффект использования яркого искусственного освещения в течение нескольких часов в день для лечения зимней депрессии.
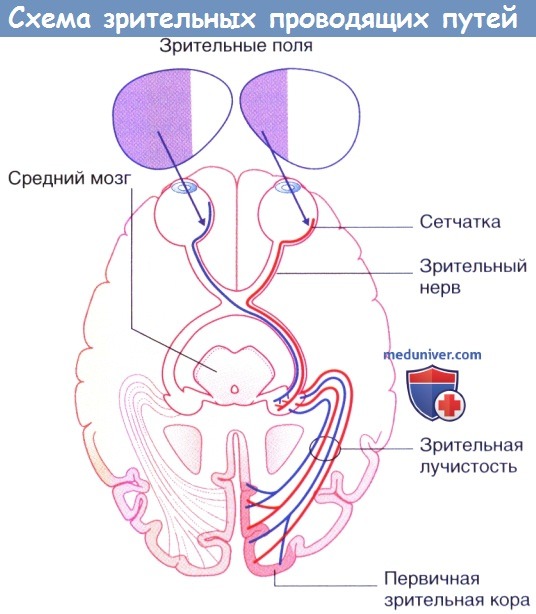
Схема зрительных проводящих путей.
Два зрительных поля (правого и левого глаза) представлены раздельно без наложения.
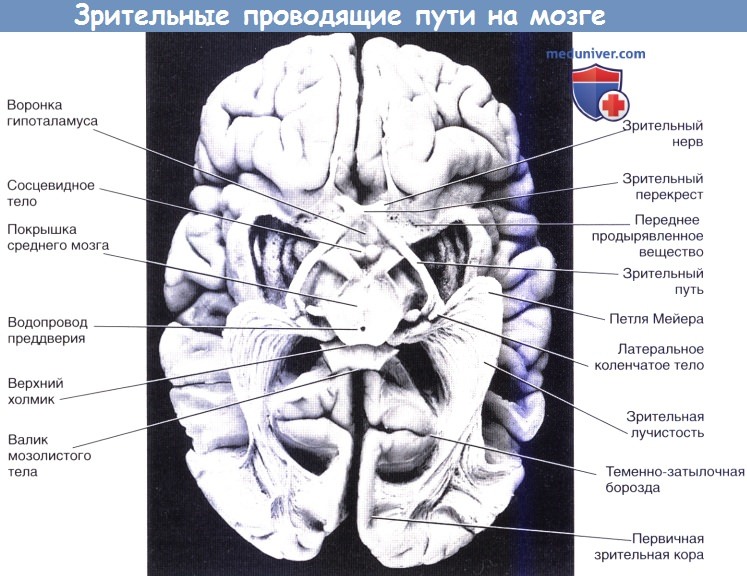
На срезе мозга показаны зрительные проводящие пути, вид снизу.
Каждый зрительный путь огибает средний мозг и разделяется на медиальный и латеральный корешки:
1. Медиальный корешок зрительного тракта. Медиальный корешок содержит 10 % волокон зрительного нерва. Он входит в средний мозг с боковой стороны и включает четыре различных группы волокон.
— Часть волокон, особенно от М-клеток сетчатки, входит в верхний холмик и обеспечивает автоматический анализ информации, как, например, при чтении этой страницы.
— Часть волокон проходит через верхний холмик к подушке таламуса; они составляют фрагмент экстраколенчатого проводящего пути к зрительной коре больших полушарий.
— Часть волокон входит в предпокрышечное ядро и участвует в зрачковом световом рефлексе.
— Часть волокон достигает мелкоклеточной ретикулярной формации, где они выполняют функцию возбуждения.
2. Латеральный корешок зрительного тракта и латеральное коленчатое тело. Латеральный корешок зрительного тракта оканчивается в латеральном коленчатом теле (ЛК’Г) таламуса. ЛКТ образовано шестью клеточными пластинками, три из которых состоят из пересеченных волокон, а три — из непересеченных (слои 2, 3, 5; «U-235»). Две наиболее глубоких пластинки (одна с пересеченными, другая — с непересеченными волокнами) образованы крупными клетками, к ним подходят аксоны ганглиозных клеток М-типа, отвечающих за регистрацию движений (расположение в пространстве, скорость, и направление). К четырем другим мелкоклеточным пластинкам подходят аксоны от Р-клеток, отвечающих за детальное зрение (цвет и детализация изображения).
Нервные цепочки ЛКТ напоминают таковые в других релейных таламических ядрах и включают тормозные (γ-аминомасляная кислота, ГАМК) терминали, отходящие от вставочных нейронов и от таламического ретикулярного ядра. (Часть ретикулярного ядра, взаимодействующая с ЛКТ, носит название околоколенчатое ядро.) Корково-коленчатые волокна начинаются в первичной зрительной коре и образуют контакты с дистальными отделами дендритов релейных клеток, а также с тормозными вставочными нейронами. Количество синапсов корковых нейронов с релейными клетками в два раза превышает количество контактов ганглиозных клеток сетчатки.
Корковая стимуляция обычно усиливает реакцию релейных клеток в ответ на импульсы, полученные от сетчатки. Вероятно, но не доказано, что их функция заключается в селективном усилении различных аспектов зрительного восприятия, таких как поиск объекта известной формы или цвета. При функциональной магнитно-резонансной томографии (фМРТ) можно обнаружить области повышенной нейрональной активности головного мозга. При фМРТ было установлено, что если доброволец ожидает увидеть на экране интересующий объект, метаболическая активность в ЛКТ увеличивается до появления стимула.
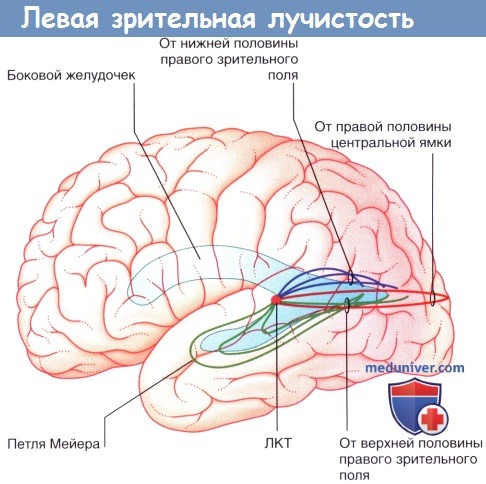
Левая зрительная лучистость. ЛКТ—Латеральное коленчатое тело.
б) Коленчато-шпорный путь и первичная зрительная кора. Зрительная лучистость (коленчато-шпорный путь) имеет важное клиническое значение, поскольку поражение этой области часто происходит при тромбозе сосудов или опухолевом процессе в заднем отделе полушария головного мозга. Проводящий путь проходит от латерального коленчатого тела к первичной зрительной коре.
Строение зрительной лучистости показано на рисунках ниже. Волокна, лежащие в нижней половине первичной зрительной коры, направляются кпереди в височную долю в виде петли Мейера, а затем поворачивают кзади и сопровождают волокна от верхней половины. Путь входит в зачечевицеобраз-ную часть внутренней капсулы и располагается в белом веществе под латеральной височной корой. Он прилежит к заднему рогу бокового желудочка до поворота в медиальном направлении и прохождения в затылочную кору.
Первичная зрительная кора занимает стенки шпорной борозды по всей ее длине (глубина борозды—10 мм). Она прилежит также к медиальной поверхности полушария на 5 мм выше и ниже борозды и к затылочному полюсу мозга на расстояние 10 мм. Общая площадь составляет 28 см2. При ранней аутопсии кору легко определяют по тонкому пучку белого вещества (зрительная полоска Дженнари) в толще серого вещества, что объясняет альтернативное название — стриарная, или полосатая, кора. Левый и правый глаза представлены в коре чередующимися полосами,называемые глазными доминантными колонками.
1. Ретинотопическая карта. Зрительное поле противоположной стороны проецируется в виде перевернутого изображения. Плоскость шпорной борозды расположена горизонтально. Сетчатка расположена спереди назад со значительно более широко представленной центральной ямкой в задней половине шпорной коры.
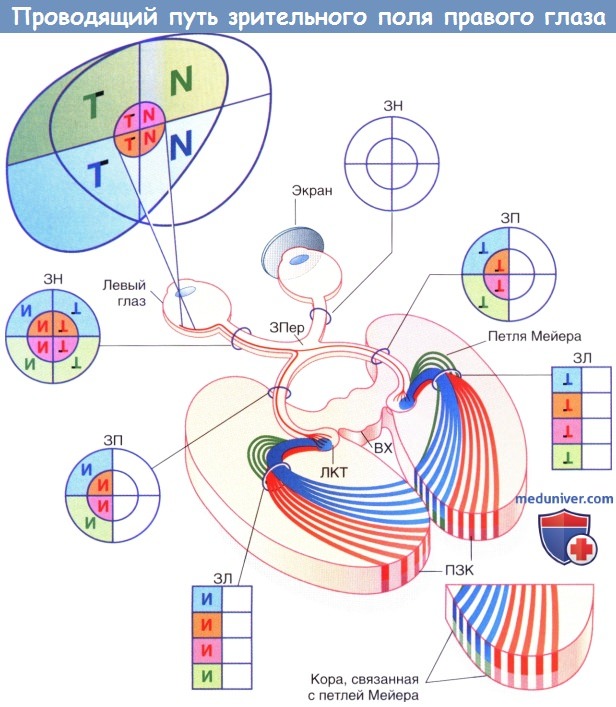
Проводящий путь от зрительного поля правого глаза к первичной зрительной коре.
Т указывает на височную (темпоральную) половину зрительного поля. N обозначает назальную (внутреннюю) половину левого зрительного поля.
В левой сетчатке и зрительном нерве (ЗН) полученное изображение представлено зеркально отраженным и перевернутым.
Правая сетчатка и зрительный нерв неактивны, так как этот глаз экранирован.
В зрительном перекресте (ЗПер) аксоны, образующие назальную половину левого зрительного нерва, пересекают среднюю линию и образуют медиальную половину правого зрительного пути (ЗП).
Волокна, расположенные в латеральной половине нерва, проходят в латеральной половине левого зрительного пути.
Каждая группа волокон образует контакты с соответствующим латеральным коленчатым телом (ЛКТ).
Зрительные лучистости (ЗЛ) имеют веерообразное строение; их аксоны, передающие информацию от центральных ямок, вначале располагаются в центре пучка.
По мере их приближения к затылочному полюсу аксоны центральной ямки (красного цвета) в обоих полушариях смещаются кзади и входят в задний отдел первичной зрительной коры (ПЗК).
Обратите внимание на тип прохождения аксонов к коре с обеих сторон (полосами).
Интервалы между ними имеют одинаковую ширину и содержат волокна, идущие к зрительной коре и образующие зрительное поле правого глаза.
ВХ—верхний холмик.
2. Поражения зрительных проводящих путей. Исследование зрительных путей имеет следующие особенности:
• Пациент может не подозревать о слепоте весьма значительной степени—в некоторых случаях даже о гемианопсии.
• Значительные зрительные нарушения можно выявить при обычном противопоставлении, как указано далее. Пациент закрывает попеременно каждый глаз и фокусирует взгляд на носу врача. Врач, сидя напротив, смотрит пациенту в глаз, внося в поле зрения одну или другую руку с различных сторон, покачивая указательным пальцем.
• В слепой зоне пациент не видит темноту—пациент вообще ничего не видит.
— Зрительные дефекты описывают с точки зрения пациента относительно зрительных полей. Для упрощения схемы представления зрительных полей, отмеченных цифрами от 1 до 9, черный цвет используют для обозначения области, в которой зрение отсутствует. На самом деле на схемах зрительных полей указывают области с сохранным зрением (в противоположность представленным графически) Слепое пятно не обозначают, но оно должно располагаться темпоральнее (латеральнее) центральной точки фиксации взгляда.]
• Возможные места повреждения зрительных проводящих путей показаны на рисунке ниже. Проявления соответствуют представленным номерам в ниже.
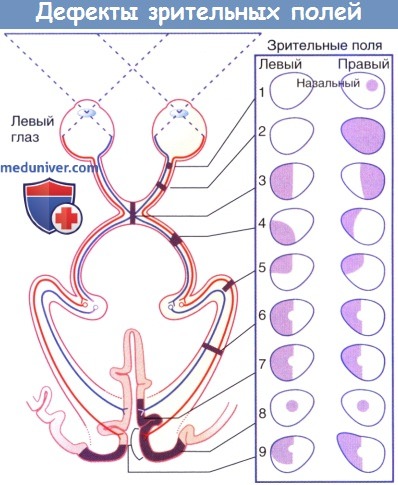
Дефекты зрительных полей, возникающие при различных поражениях зрительных проводящих путей:
1. Частичное, зрительного нерва — Ипсилатеральная скотома
2. Полное, зрительного нерва — Слепота на этот глаз
3. Зрительный перекрест — Битемпоральная гемианопсия
4. Зрительный путь — Контралатеральная гомонимная геминанопсия
5. Петля Мейера — Контралатеральная гомонимная верхняя квадрантанопсия
6. Зрительная лучистость — Контралатеральная гомонимная гемианопсия
7. Зрительная кора — Контралатеральная гомонимная гемианопсия
8. Макулярная кора, двустороннее — Двусторонние центральные скотомы
9. Задние отделы зрительной коры — Височная серповидная контралатеральная гомонимная гемианопсия
Примечания по приведенным выше поражениям:
1. Эксцентрические поражения зрительного нерва приводят к образованию скотом в назальном или височном поле зрения пораженного глаза. При развитии скотомы у молодых взрослых всегда следует подозревать рассеянный склероз.
2. Полное поражение зрительного нерва может возникать при черепно-мозговой травме.
3. Сдавление середины перекреста чаще всего бывает вызвано аденомой (доброкачественной опухолью) гипофиза.
4. Поражения зрительного пути встречают редко. Несмотря на то, что выпадают гомонимные (односторонние) зрительные поля, наружная, неприкрытая половина зрительного пути, поражается чаще, чем внутренняя половина, поэтому гемианопсию обозначают как несимметричную.
5. Избирательное поражение петли Мейера встречают при опухолях височной доли.
6. Поражение зрительной лучистости встречают при опухолях височной, теменной или затылочной долей. Зрительные поля обоих глаз обычно выпадают в одинаковой степени (симметрично), и желтое пятно остается интактным. Опухоли, инфильтрирующие лучистость снизу, приводят к дефекту в нижнем квадранте. Основной пучок лучистости расположен в зачечевицеобразном отделе внутренней капсулы и часто поражается при отеке, сопровождающем кровотечение из ветви средней мозговой артерии (классический инсульт).
7. Тромбоз задней мозговой артерии сопровождается гомонимной гемианопсией. Пробелы в зрительном поле № 7 указывают на сохранность желтого пятна (макулы). Сохранность макулярных половин полей зрения непостоянна и часто обусловлена двойным кровоснабжением затылочного полюса из средней и задней мозговых артерий.
8. Двусторонние центральные скотомы наиболее часто развиваются при падении на затылок с ушибом мозга в затылочном отделе.
9. Височную серповидную гомонимную гемианопсию встречают при повреждении затылочной коры с сохранением наиболее переднего ее отдела—области, к которой подходят зрительные лучистости, идущие от назального отдела сетчатки (назальная половина сетчатки «видит» височное зрительное поле).
в) Резюме. В процессе эмбриогенеза сетчатка развивается из наружного выпячивания диэнцефалона. Эмбриональный зрительный бокал состоит из наружного пигментного слоя и внутреннего оптического слоя, между которыми расположено внутрисетчаточное (интраретинальное) пространство. Оптический слой образован тремя типами радиально расположенных нейронов (фоторецепторами, биполярными клетками и ганглиозными клетками) и двумя типами тангенциально (по касательной линии) расположенных клеток (горизонтальными и амакриновыми). Во всех отделах кроме центральной ямки свет должен пройти через другие слои сетчатки, чтобы попасть на фоторецепторы.
Две трети зрительного поля — бинокулярные, наружная 1/6 с каждой стороны—монокулярная. Зрительные дефекты описывают относительно зрительных полей.
Палочковые фоторецепторы функционируют при сумеречном свете и отсутствуют в центральной ямке. Колбочки наиболее многочисленны в центральной ямке; они отвечают за распознавание формы и имеют три типа чувствительности к цвету. Ганглиозные клетки имеют концентрическую реакцию на возбуждения по типу «от центра к периферии» с цветовым противодействием. М-ганглиозные клетки—относительно крупные и отвечают за регистрацию движений, их аксоны направляются к двум крупноклеточным слоям в ЛКТ. Функция Р-ганглиозных клеток—распознавание отдельных свойств изображения и цвета, они взаимодействуют с четырьмя мелкоклеточными слоями ЛКТ.
ЛКТ — двустороннее образование, получающее информацию от противоположной назальной половины сетчатки (через зрительный перекрест), а также от ипсилатеральной височной половины сетчатки. Обе группы аксонов проходят через зрительный тракт, от которого также отходят коллатерали к среднему мозгу для образования низших зрительных рефлексов.
Зрительная лучистость (коленчато-шпорный тракт) начинается от М- и Р-клеток ЛКТ и огибает боковой желудочек, достигая первичной зрительной коры в стенках шпорной борозды.
Разнообразные дефекты зрительных полей возникают вследствие повреждения любого из пяти главных отделов зрительных проводящих путей (зрительного нерва, зрительного перекреста, зрительного тракта, зрительной лучистости и зрительной коры).
— Вернуться в оглавление раздела «Неврология.»
Редактор: Искандер Милевски. Дата публикации: 21.11.2018
Источник
Запрос «Зрение» перенаправляется сюда; об альбоме певицы Линды см. Зрение (альбом).
Проводящие пути зрительного анализатора:
1 — Левая половина зрительного поля,
2 — Правая половина зрительного поля,
3 — Глаз,
4 — Сетчатка,
5 — Зрительные нервы,
6 — Глазодвигательный нерв,
7 — Хиазма,
8 — Зрительный тракт,
9 — Латеральное коленчатое тело,
10 — Верхние бугры четверохолмия,
11 — Неспецифический зрительный путь,
12 — Зрительная кора головного мозга.
Зри́тельная систе́ма (зри́тельный анализа́тор, о́рган зре́ния) — бинокулярная (стереоскопическая) оптическая система биологической природы, эволюционно возникшая у животных и способная воспринимать электромагнитное излучение видимого спектра (свет), создавая ощущение положения предметов в пространстве. Зрительная система обеспечивает функцию зрения.
Нормальным раздражителем органа зрения является свет. Под влиянием света в палочках, колбочках (см. ниже) и светочувствительных ганглионарных клетках происходит распад зрительных пигментов (родопсина, йодопсина и меланопсина). Палочки функционируют при свете слабой интенсивности, в сумерках; зрительные ощущения, получаемые при этом, бесцветны. Колбочки функционируют днём и при ярком освещении; их функция определяет ощущение цветности.
Человек и многие другие животные обладают бинокулярным зрением, обеспечивающим возможность воспринимать объёмное изображение. Большинство дневных животных также обладает способностью различать отдельные цвета солнечного света (цветовое зрение).
Анатомия[править | править код]
Зрительная система (зрительный анализатор) у млекопитающих включает следующие анатомические образования:
- периферический парный орган зрения — глаз (с его воспринимающими свет фоторецепторами — палочками, колбочками и светочувствительными ганглионарными клетками сетчатки);
- нервные структуры и образования ЦНС: зрительные нервы, хиазма, зрительный тракт, зрительные пути — II пара черепных нервов, глазодвигательный нерв — III пара, блоковый нерв — IV пара и отводящий нерв — VI пара;
- латеральное коленчатое тело промежуточного мозга (с подкорковыми зрительными центрами), передние бугры четверохолмия среднего мозга (первичные зрительные центры);
- подкорковые (и стволовые) и корковые зрительные центры: латеральное коленчатое тело и подушки зрительного бугра, верхние холмики крыши среднего мозга (четверохолмия) и зрительная кора.
Кровоснабжение[править | править код]
Кровоснабжение осуществляется из бассейна внутренней сонной артерии по глазным артериям. Сетчатка кровоснабжается центральной артерией сетчатки, которая проникает в глаз в составе (в толще) зрительного нерва, а также получает кровь из сосудистой оболочки глаза. Слёзная железа получает кровь из слёзной артерии. Мышцы глаза — из одноимённых артерий.
Венозная кровь от различных структур и образований глазного яблока оттекает по одноимённым венам, которые сливаясь образуют верхнюю и нижнюю глазные вены. От сетчатки кровь оттекает в центральную вену сетчатки, из сосудистой оболочки — в четыре вены, впадающие в глазные вены.
Дополнительные структуры глаза[править | править код]
Мышцы[править | править код]
Внешние мышцы глазного яблока (6)[1]:
- наружная прямая мышца глаза;
- внутренняя прямая мышца глаза;
- нижняя прямая мышца глаза;
- верхняя прямая мышца глаза;
- нижняя косая мышца глаза;
- верхняя косая мышца глаза.
Орбиты[править | править код]
Орби́та (глазни́ца) — парная полость в черепе, представляющая собой пирамидальную впадину, обладающую основанием, вершиной и четырьмя стенками. Содержит глазное яблоко с его придатками[2].
Слёзный аппарат[править | править код]
Слёзный аппарат состоит из слёзных желез и системы слёзных путей. Выделяет и транспортирует к глазному яблоку слёзную жидкость (слезу), вырабатываемую гардеровыми или слёзными железами для смачивания и очищения поверхности глаза у земноводных, пресмыкающих, птиц и млекопитающих. Эта жидкость прозрачная, слегка опалесцирует, имеет слабощелочную реакцию (норма рН слёзной жидкости: 7,3…7,5). Вырабатываемая слёзная жидкость по выводным канальцам (лат. ductuli excretorii) поступает в конъюнктивальный мешок (лат. saccus conjunctivae) и скапливается в нём, а оттуда движением век переносится на роговицу. После этого слёзная жидкость через слёзоотводящие пути — слёзное озеро (лат. lacus lacrimalis), слёзные канальцы (лат. canaliculi lacrimales), слёзный мешок (лат. saccus lacrimalis) и слёзно-носовой проток (лат. ductus nasolacrimalis) — уходит в нижний носовой ход[3][4].
Глаза[править | править код]
Основная статья: Глаз
У животных и человека органами зрения являются глаза. Высокоорганизованными (способными создавать изображения предметов и обеспечивать предметное зрение) глазами обладают, помимо позвоночных, головоногие моллюски и многие членистоногие, а также отдельные представители других типов животных — книдарий, кольчатых червей, плоских червей.[5] Фасеточные глаза насекомых имеют принципиально отличное строение по сравнению с камерными глазами позвоночных и головоногих, однако связаны с ними постепенными переходами сравнительно-морфологического ряда.
Альтернативные способы ориентирования в пространстве[править | править код]
Существуют сходные по функции со зрением другие сенсорные системы, применяемые для ориентирования в пространстве, например, ультразвуковая эхолокация летучих мышей и китообразных, позволяющая им обнаруживать мельчайшие объекты, электролокация некоторых рыб и утконоса, тепловая локация гремучих змей.
Также для ориентирования в пространстве применяются обоняние (наиболее характерен в этом смысле язык у змей, хотя так же широко известны в качестве примера ориентации по запаху у собаки), слух (боковая линия у рыб), и тактильные ощущения (восприятие давления и температуры, ощупывание).
Эволюция зрительной системы[править | править код]
Беспозвоночные[править | править код]
Как установлено с помощью методов генетической трансформации, гены eyeless дрозофилы и Small eye мыши, имеющие высокую степень гомологии, контролируют развитие глаза: при создании генноинженерной конструкции, с помощью которой вызывалась экспрессия гена мыши в различных имагинальных дисках мухи, у мухи появлялись эктопические фасеточные глаза на ногах, крыльях и других участках тела.[6] В целом в развитие глаза вовлечено несколько тысяч генов, однако один-единственный «пусковой ген» («мастер-ген») осуществляет запуск всей этой генной сети. То, что этот ген сохранил свою функцию у столь далёких групп, как насекомые и позвоночные, может свидетельствовать об общем происхождении глаз всех двустороннесимметричных животных.
Позвоночные[править | править код]
Глазные бокалы позвоночных формируются как выросты промежуточного мозга, а первичный центр обработки зрительной информации находится в среднем мозге.
Млекопитающие[править | править код]
Предполагается, что в течение мезозойского периода ранние млекопитающие занимали подчинённое по отношению к «царствующим рептилиям» (особенно динозаврам, преимущественно занимавшим экологические ниши крупных хищников и травоядных) положение, имели мелкие размеры и сумеречный образ жизни. В таких условиях зрение для ориентации в пространстве становится второстепенным по отношению к обонянию и слуху. Химические чувства, которые и сейчас остаются для нас эмоционально окрашенными, обслуживаются передним мозгом и лимбической системой. Предполагается, что передний мозг в этих условиях приобретает большее значение. Когда «царствующие» рептилии исчезли в конце мезозоя, более широкие эволюционные возможности открылись для «угнетённых» млекопитающих. Они заселили все возможные экологические ниши освободившегося мира, зрение для некоторых отрядов снова стало наиболее важным из всех чувств. Однако формирующиеся заново зрительные пути направились к наиболее важной части мозга — переднему мозгу, расширяющемуся и формирующему характерные для млекопитающих крупные полушария. Ретино-тектальный путь остается пережитком старого зрительного пути, а ретино-геникуло-стриарный путь быстро становится наиболее важным путём передачи зрительной информации в мозг.
Зрительная система у разных таксономических групп[править | править код]
Беспозвоночные[править | править код]
У беспозвоночных встречаются очень разнообразные по типу строения и зрительным возможностям глаза и глазки — одноклеточные и многоклеточные, прямые и обращённые (инвертированные), паренхимные и эпителиальные, простые и сложные.
У членистоногих часто присутствует несколько простых глаз (иногда непарный простой глазок — например, науплиальный глаз ракообразных) или пара сложных фасеточных глаз. Среди членистоногих некоторые виды имеют и простые, и сложные глаза: так, у ос два сложных глаза и три простых глаза (глазка). У скорпионов 3—6 пар глаз (1 пара — главные, или медиальные, остальные — боковые), у щитня — 3. В эволюции фасеточные глаза произошли путём слияния простых глазков. Близкие по строению к простому глазу, глаза мечехвостов и скорпионов, видимо, возникли из сложных глаз трилобитообразных предков путём слияния их элементов (Беклемишев, 1964).
Простейшие[править | править код]
Некоторые простейшие имеют слабодифференцированные органоиды светового восприятия (например, стигма у эвглены зелёной).
Насекомые[править | править код]
Глаза насекомых имеют фасеточное строение. Разные виды по-разному воспринимают цвета, но в целом большинство насекомых хорошо различают не только лучи спектра, видимые человеком, но и ближний ультрафиолет. Это зависит, помимо генетических факторов (строение рецепторов), и от меньшего поглощения УФ-света — из-за меньшего его пути в оптической системе глаза. Например, пчёлы видят ультрафиолетовый рисунок на цветке.
Позвоночные[править | править код]
Зрительная система рептилий, птиц и некоторых рыб[править | править код]
Установлено, что рептилии, птицы и некоторые рыбы имеют более широкую область ощущаемого оптического излучения. Они воспринимают ближний ультрафиолет (300—380 нм), синюю, зелёную и красную часть спектра. У некоторых земноводных, например, гребенчатого тритона, как показал Р. Маттей в 1925 году, зрение способно восстанавливаться после перерезания зрительного нерва[7].
Зрительный аппарат птиц обладает особенностями, не сохранившимися в зрении человека. Так, в рецепторах птиц имеются микросферы, содержащие липиды и каротиноиды. Считается, что эти микросферы — бесцветные, а также окрашенные в жёлтый или оранжевый цвет — выполняют функцию специфических светофильтров, формирующих «кривую видности».
У многих птиц их бинокулярное зрение из-за специфического расположения глаз не даёт такого большого поля стереоскопического зрения, как у человека.
Зрение млекопитающих[править | править код]
Мутация, некогда реализованная у одного из прапредков млекопитающих и закрепившаяся во всём классе, сократила число видов цветовых рецепторов колбочек до двух. Полагают, что предки млекопитающих — мелкие грызуны — вели ночной образ жизни и компенсировали эту потерю значительным развитием сумеречного зрения (с помощью рецепторов — палочек).
Позже, однако, у приматов (в том числе человека) другая мутация вызвала появление третьего типа колбочек — цветовых рецепторов. Это было вызвано расширением экологической ниши млекопитающих, переходом части видов к дневному образу жизни, в том числе на деревьях. Мутация была вызвана появлением изменённой копии гена, отвечающего за восприятие средней, зелёночувствительной области спектра. Она обеспечила лучшее распознавание объектов «дневного мира» — плодов, цветов, листьев.
Глаз человека состоит из глазного яблока и зрительного нерва с его оболочками.
У человека и позвоночных имеется по два глаза, расположенных в глазных впадинах черепа.
Стереоскопическое зрение[править | править код]
У многих видов, образ жизни которых требует хорошей оценки расстояния до объекта, глаза смотрят скорее вперёд, нежели в стороны. Так, у горных баранов, леопардов, обезьян обеспечивается лучшее стереоскопическое зрение, которое помогает оценивать расстояние перед прыжком. Человек также имеет хорошее стереоскопическое зрение (см. ниже, раздел Бинокулярное и стереоскопическое зрение).
Альтернативный механизм оценки расстояния до объекта реализован у некоторых птиц, глаза которых расположены по разным сторонам головы, а поле объёмного зрения невелико. Так, куры совершают постоянные колебательные движения головой, при этом изображение на сетчатке быстро смещается, обратно пропорционально расстоянию до объекта. Мозг обрабатывает сигнал, что позволяет поймать мелкую добычу клювом с высокой точностью.
Глаза каждого человека внешне кажутся идентичными, но всё же функционально несколько различны, поэтому выделяют ведущий и ведомый глаз.
Определение ведущего глаза важно для охотников, видеооператоров и лиц других профессий. Если посмотреть через отверстие в непрозрачном экране (дырочка в листе бумаги на расстоянии 20—30 см) на отдалённый предмет, а затем, не смещая голову, поочерёдно закрыть правый и левый глаз, то для ведущего глаза изображение не сместится.
Физиология зрения человека[править | править код]
Из-за большого числа этапов процесса зрительного восприятия его отдельные характеристики рассматриваются с точки зрения разных наук — оптики, психологии, физиологии, химии.
Бинокулярное зрение у человека, как и у других млекопитающих, а также птиц и рыб, обеспечивается наличием двух глаз, информация от которых обрабатывается сначала раздельно и параллельно, а затем синтезируется в мозгу в зрительный образ. У далёких филогенетических предшественников человека глаза были расположены латерально, их зрительные поля не перекрывались и каждый глаз был связан только с противоположным полушарием мозга — контралатерально. В процессе эволюции у некоторых позвоночных, в том числе и у предков человека в связи с приобретением стереоскопического зрения, глаза переместились вперёд. Это привело к перекрытию левого и правого зрительных полей и к появлению новых ипсилатеральных связей: левый глаз — левое полушарие, правый глаз — правое. Таким образом появилась возможность иметь в одном месте зрительную информацию от левого и правого глаза, для их сопоставления и измерения глубины.
Ипсилатеральные связи эволюционно более молодые, чем контралатеральные. В ходе развития стереоскопичности зрения по мере перехода от животных с латерально направленными зрительными осями к животным с фронтальной ориентацией глаз доля ипси-волокон растёт (таблица)[8].
| Вид животного | Отношение количества неперекрёстных к числу перекрёстных волокон |
|---|---|
| Овца | 1:9 |
| Лошадь | 1:8 |
| Собака | 1:4,5 |
| Опоссум | 1:4 |
| Морская свинка | 1:3 |
| Кошка | 1:3 |
| Хорёк | 1:3 |
| Макака | 1:1,5 |
| Человек | 1:2; 1:1,5; 1:1[9] |
Большинство особенностей бинокулярного зрения человека обусловлено характеристиками нейронов и нейронных связей. Методами нейрофизиологии показано, что декодировать глубину изображения, заданную на сетчатках набором диспаратностей, начинают бинокулярные нейроны первичной зрительной коры. Было показано, что самое важное требование для осуществления стереоскопического зрения — это различия в образах на сетчатке двух глаз.[10]
Благодаря тому, что поля зрения обоих глаз человека и высших приматов в значительной мере пересекаются, человек способен лучше, чем многие млекопитающие, определять внешний вид и расстояние (этому способствует также механизм аккомодации) до близких предметов в основном за счёт эффекта стереоскопичности зрения. Стереоскопический эффект сохраняется на дистанции приблизительно 0,1—100 м. У человека пространственно-зрительные способности и объёмное воображение тесно связаны со стереоскопией и ипси-связями.
Примечания[править | править код]
- ↑ Ханц Фениш. Карманный атлас анатомии человека. Минск: Вышэйшая школа, 1996 г.
- ↑ Сапин М. Р., Брыксина З. Г. — Анатомия человека //Просвещение, 1995 г.
- ↑ Гистология, цитология и эмбриология, 2004, с. 362.
- ↑ Слёзный аппарат глаза. Дата обращения 30 марта 2019. Архивировано 8 августа 2018 года.
- ↑ Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных. Т. 2. — М., Наука, 1964. — с. 143—159.
- ↑ glava 14.1.p65 Архивировано 19 апреля 2009 года.
- ↑ R. Matthey. Récupération de la vue après résection des nerfs optiques chez le Triton (фр.) // Comptes rendus des séances de la Société de biologie et de ses filiales : magazine. — 1925. — Vol. 93. — P. 904—906.
- ↑ Блинков С. М., Глезер И. И. Мозг человека в цифрах и таблицах. — Л., 1964. — 180 с.
- ↑ Данные разных авторов.
- ↑ Bishop P. O. (1981) Neural mechanisms for binocular depth discrimination. In: Advances in Physiological Sciences. Sensory Functions (Eds. Grastian E., Molnar P.), v. 16, p. 441—449.
См. также[править | править код]
- Зрение человека
- Рецептивное поле
- Сенсорная система
- Человек разумный
Литература[править | править код]
- Зрение // Энциклопедический словарь Брокгауза и Ефрона : в 86 т. (82 т. и 4 доп.). — СПб., 1890—1907.
- А. Нагель «Аномалии, рефракции и аккомодации глаза» (перевод с немецкого доктора В. Добровольского, 1881);
- Т. Лонгмор, «Руководство к исследованию зрения для военных врачей» (переработано доктором медицины Лаврентьевым, 1894);
- А. Imbert, «Les anomalies de la vision» (1889);
- Дж. Грегг, «Опыты со зрением» (1970).
- Гистология, цитология и эмбриология. 6-е изд / Под ред. Ю. И. Афанасьева, С. Л. Кузнецова, H. А. Юриной. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
Ссылки[править | править код]
- Зрительный аппарат человека
- Болезни глаз
- Зрительная система человека
Источник

