
В состав сетчатки входят амакриновые клетки
Амакриновые клетки. Ганглиозные клетки сетчатки
В настоящее время морфологическими или гистохимическими методами идентифицированы около 30 типов амакриновых клеток. Функции некоторых из них охарактеризованы, и все они отличаются друг от друга. Один тип амакриновых клеток является частью прямого пути палочкового зрения, т.е. от палочки к биполярным клеткам, затем к амакриновым и, наконец, к ганглиозным клеткам.
Амакриновые клетки другого типа активно реагируют в начале непрерывного зрительного сигнала, но ответ быстро исчезает. Некоторые амакриновые клетки, наоборот, мощно реагируют на выключение зрительного сигнала, но их ответ также быстро прекращается.
Амакриновые клетки еще одного типа реагируют и на включение, и на выключение света, просто сигнализируя об изменении освещения, независимо от его направления. Есть амакриновые клетки, реагирующие на движение пятна света по сетчатке в определенном направлении; о таких клетках говорят, что они чувствительны к направлению.
В известном смысле большинство амакриновых клеток являются вставочными нейронами, которые помогают анализировать зрительные сигналы, прежде чем они покинут сетчатку.
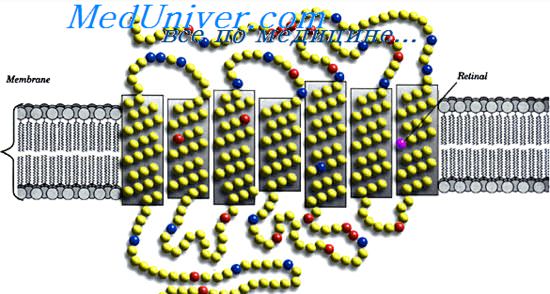
Ганглиозные клетки
Каждая сетчатка содержит около 100 млн палочек и 3 млн колбочек; однако количество ганглиозных клеток — лишь около 1,6 млн. Таким образом, в среднем 60 палочек и 2 колбочки конвергируют на каждую ганглиозную клетку и волокно зрительного нерва, идущее от ганглиозной клетки к мозгу.
Однако существуют большие различия между периферической и центральной областями сетчатки. По мере приближения к ямке все меньше палочек и колбочек конвергируют на каждое зрительное волокно; кроме того, и палочки, и колбочки становятся тоньше. Эти эффекты постепенно увеличивают остроту зрения в центральной сетчатке. В самом центре — в области центральной ямки — есть только тонкие колбочки (примерно 35000) и совсем нет палочек. К тому же, как показано справа на рисунке, количество волокон зрительного нерва, выходящих из этой части сетчатки, почти равно числу колбочек. Это объясняет высокую степень остроты зрения в центральной сетчатке по сравнению с гораздо меньшей остротой на периферии.
Другим различием между периферией и центром сетчатки является гораздо более высокая чувствительность периферической сетчатки к слабому свету. Отчасти это объясняется тем, что чувствительность палочек в 30-300 раз выше, чем колбочек. Однако эффект значительно усиливается в связи с конвергенцией примерно 200 палочек на одну ганглиозную клетку в периферических областях сетчатки, поэтому сигналы от палочек суммируются, дополнительно усиливая интенсивность стимуляции периферических ганглиозных клеток и исходящих от них волокон зрительных нервов.
Различают три группы ганглиозных клеток, которые обозначают как W-, Х- и Y-клетки. Каждая группа выполняет свою функцию.
Передача палочкового зрения W-клетками. W-клетки составляют примерно 40% общего числа ганглиозных клеток. Они маленькие (диаметром около 10 мкм) и передают сигналы по соответствующим им волокнам зрительного нерва с низкой скоростью, равной около 8 м/сек. Эти ганглиозные клетки возбуждаются в основном от палочек, передающих к ним сигналы через малые биполярные и амакриновые клетки. W-клетки имеют широкие рецептивные поля в сетчатке, т.к. их дендриты широко распространяются во внутреннем слое сетчатки, получая сигналы от обширных областей.
На основании гистологических и физиологических экспериментов показано, что W-клетки, по-видимому, особенно чувствительны к восприятию направленного движения в поле зрения и очень важны для нашего грубого палочкового зрения в условиях темноты.
Передача зрительного образа и цвета Х-клетками. Подавляющее большинство ганглиозных клеток (55%) являются Х-клетками. Они имеют средний диаметр (в пределах от 10 до 15 мкм) и передают сигналы по своим волокнам зрительного нерва со скоростью около 14 м/сек.
Рецептивные поля Х-клеток небольшие, поскольку их дендриты не имеют широкого распространения в сетчатке. В связи с этим сигналы Х-клеток отражают дискретные участки сетчатки. Следовательно, в основном через Х-клетки передаются тонкие детали зрительного образа. Кроме того, поскольку каждая Х-клетка получает сигналы, по крайней мере, от одной колбочки, эти клетки, вероятно, отвечают за все цветовое зрение.
Функция Y-клеток — передача информации о мгновенных изменениях в зрительном образе. Y-клетки — самые большие из всех ганглиозных клеток (диаметром до 35 мкм). Они проводят сигналы к мозгу со скоростью 50 м/сек и выше. Среди ганглиозных клеток они самые малочисленные (около 5% общего количества) и имеют разветвленные дендриты, следовательно, собирают сигналы от обширных областей сетчатки.
Как многие из амакриновых клеток, Y-клетки реагируют на быстрые изменения в зрительном образе (быстрые движения или быстрые изменения освещения), посылая импульсные разряды, длительность которых составляет лишь доли секунды. Эти ганглиозные клетки, вероятно, почти немедленно информируют центральную нервную систему о появлении любого нового зрительного явления в поле зрения, но без высокой точности его локализации, обеспечивая лишь соответствующие сигналы, заставляющие глаза двигаться по направлению к возбуждающему объекту.
— Также рекомендуем «Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке»
Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»:
1. Каскад усиления в сетчатке. Фотохимия цветового зрения
2. Световая и темновая адаптация. Механизмы световой и темновой адаптации
3. Острота зрения. Определение расстояния до объекта глазами
4. Слепота на отдельные цвета. Функция нейронов сетчатки
5. Зрительный путь от колбочек. Нейромедиаторы нейронов сетчатки
6. Функция горизонтальных клеток сетчатки. Возбуждение и торможение биполярных клеток
7. Амакриновые клетки. Ганглиозные клетки сетчатки
8. Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке
9. Зрительные пути. Дорсолатеральное коленчатое ядро таламуса
10. Зрительная кора. Строение первичной зрительной коры
Источник
Зрительные проводящие пути имеют важнейшее значение в клинической неврологии. Они проходят от сетчатки глаз до затылочных долей коры головного мозга. Большая протяженность путей обусловливает их особенную уязвимость для деми-елинизирующих заболеваний (рассеянный склероз), опухолей мозга или гипофиза, сосудистых поражений в бассейне средней или задней мозговых артерий или черепно-мозговых травм.
К зрительной системе относят: сетчатки, зрительные проводящие пути от сетчаток к стволу мозга и зрительной коре, а также корковые области, выполняющие высшие зрительные функции. В этой главе описаны только сетчатка и зрительные проводящие пути. Высшие зрительные функции обсуждены в главе 29.
Сечатка и зрительные нервы — части центральной нервной системы. Сетчатка эмбриона формируется из выпячивания диэнцефалона — глазного пузырька. Глазной пузырек образует инвагинацию (хрусталик) и становится двуслойным глазным бокалом.
Наружный слой глазного бокала преобразуется в пигментный эпителий зрелой сетчатки. Внутренний (оптический) слой бокала дает начало нейронам сетчатки.
На рисунке ниже показано общее топографическое строение сетчатки эмбриона. Оптический отдел образован тремя главными слоями нейронов: слоем фоторецепторов, который будет прилежать к пигментному слою клеток после резорбции внутрисетчаточного (интраретиналъного) пространства, слоем биполярных нейронов и слоем ганглиозных клеток, которые дают начало зрительному нерву и достигают таламуса и среднего мозга.
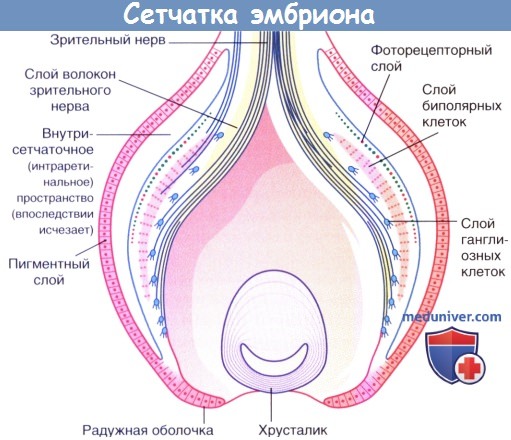
Сетчатка эмбриона.
Зеленым и красным цветом показаны палочки и колбочки соответственно.
Обратите внимание на инвертированное положение сетчатки. Свет должен пройти через слой волокон зрительного нерва, слой ганглиозных клеток и слой биполярных нейронов, чтобы достичь фоторецепторов. «Причина» расположения фоторецепторов, при котором они «максимально удалены» от источника их возбуждения (света или фотонов), обусловлена многими факторами. Во-первых, при таком расположении апикальные концы фоторецепторов (содержащие светочувствительный фотопигмент) расположены напротив пигментного слоя сетчатки, который способен поглощать любой рассеянный свет или свет, не реагирующий с фоторецепторными клетками. Во-вторых, клетки пигментного эпителия сетчатки выполняют фагоцитирующую функцию.
Светочувствительный фотопигмент палочек имеет короткий период полураспада, что требует его постоянного восполнения. Новый фотопигмент продуцируется в основании палочки и перемещается к верхушке клетки, старые апикальные компоненты сбрасываются и фагоцитируются пигментными клетками сетчатки, а белки используются заново (колбочки не сбрасывают). Наконец, фоторецепторные клетки имеют высокий уровень метаболизма и в наиболее глубоком отделе сетчатки они располагаются ближе всего к капиллярам сосудистой оболочки (лежащим под пигментным эпителием), обеспечивающим их питание.
В точке наиболее острого зрения — ямочке (фовеоле) — слои биполярных и ганглиозных клеток огибают центральную ямку (фовеа), и свет проходит к фоторецепторам с минимальным рассеянием (см. ниже «Специализация центральной ямки»). Центральная ямка зрелого глаза имеет диаметр около 1,5 мм и расположена в центре желтого пятна (macula lutea) шириной 5 мм, множество фоторецепторов которого содержат желтый пигмент. Центральная ямка — область наиболее острого зрения — расположена на зрительной оси—линии, проведен ной от центра зрительного поля глаза через центр хрусталика к центральной ямке. Для фиксации, или фовеации, объекта взгляд направляют точно на него, чтобы свет, отраженный от центра объекта, зафиксировался на центральной ямке.
Аксоны ганглиозных клеток входят в зрительный нерв через головку зрительного нерва (сосок зрительного нерва), лишенную нейронов сетчатки и образующую физиологическое слепое пятно.
Зрительные поля глаз перекрывают друг друга в двух третях общего поля зрения. Кнаружи от этого бинокулярного поля зрения с каждой стороны расположено монокулярное (височное) серповидное поле зрения. При прохождении через зрачок формируется перевернутое изображение, поэтому объекты в левой половине бинокулярного поля зрения проецируются на правую половину каждой сетчатки, а объекты в верхней части зрительного поля — на нижнюю половину. Такое расположение сохраняется на всем протяжении до зрительной коры затылочной доли.
С клинической точки зрения необходимо учитывать, что зрение—это перекрестное чувство. Зрительное поле с одной стороны зрительной оси регистрируется на зрительной коре противоположной стороны. В сущности, правая зрительная кора «видит левое поле зрения» или пространство, и наоборот. Только половина зрительной информации от каждой сетчатки пересекает зрительный перекрест по той простой причине, что другая половина уже пересекла среднюю линию.
Дефекты поля зрения, обусловленные поражением зрительных проводящих путей, всегда описывают с точки зрения пациента, т.е. в отношении полей зрения, а не в отношении топографии сетчатки.
Строение сетчатки. Помимо расположенных рядами фоторецепторных клеток, биполярных и ганглиозных клеток, показанных на рисунке ниже, в сетчатке находятся также две группы поперечно расположенных нейронов: горизонтальные клетки и амакриновые клетки. Все восемь слоев сетчатки составляют единое целое.
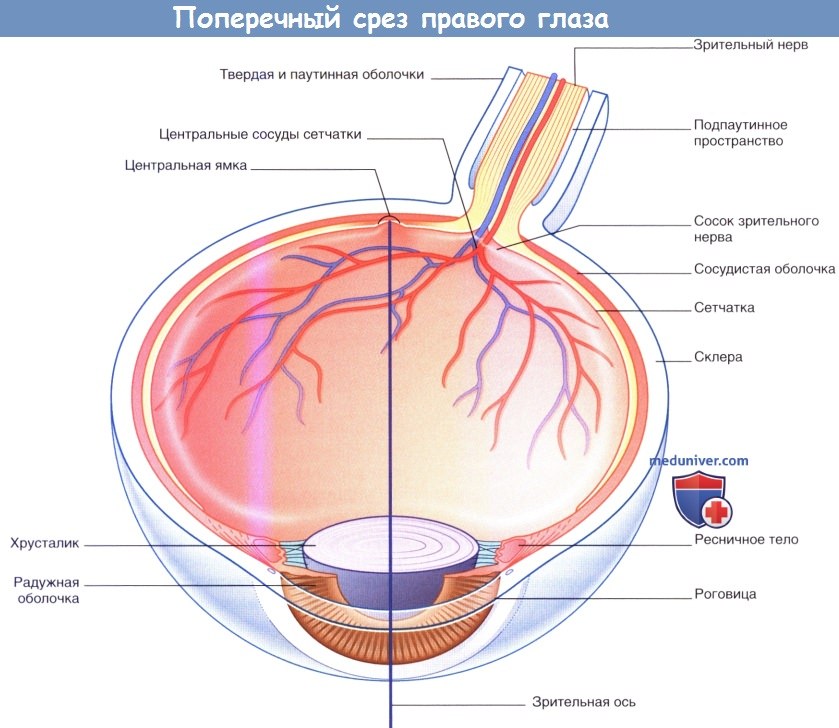
Поперечный срез правого глаза, показана зрительная ось.
Ганглиозные клетки генерируют потенциалы действия, обеспечивающие «необходимую скорость проведения» к таламусу и среднему мозгу. Расстояния между другими клетками очень короткие, поэтому для межклеточного взаимодействия бывает достаточно пассивного электрического заряда (электротонуса) или постепенных изменений мембранного потенциала клетки без образования синаптических контактов и высвобождения нейромедиатора.
1. Фоторецепторы. К фоторецепторным нейронам относят палочки и колбочки.
Палочки функционируют только при сумеречном свете и нечувствительны к цвету (электромагнитное излучение с волнами разной длины). Лишь в небольшом количестве они представлены в наружной части центральной ямки и полностью отсутствуют в ее центре. Колбочки реагируют на яркий свет, восприимчивы к цвету, форме и наиболее многочисленны в центральной ямке (в глазе человека расположено около 130 млн. фоторецепторных клеток; отношение палочек к колбочкам составляет 20:1 во всех отделах за исключением центральной ямки).
Каждая фоторецепторная клетка имеет наружный и внутренний сегменты, а также синаптическое окончание. В наружном сегменте (светочувствительной «органелле») находятся сотни мембранных дисков (у палочек) или мембранных полудисков (в колбочках), в которые упакован зрительный пигмент (родопсин — фотопигмент, поглощающий свет или фотоны и инициирующий каскад молекулярных реакций, приводящий к изменению потенциала фоторецептора и высвобождению нейромедиатора из синаптической области; этот процесс называют фотопреобразованием). Новые диски образуются во внутреннем сегменте палочек и переносятся в наружный сегмент, старые диски удаляются с апикальной области наружного сегмента. Синаптическое окончание контактирует с отростками биполярных и горизонтальных клеток в наружном ретикулярном слое.
Фоторецепторы обладают удивительным свойством гиперполяризации под действием света. В темноте натриевые (Na+) каналы открыты, образуя достаточный положительный электротонус, приводящий к высвобождению нейромедиатора (глутамата) из синаптического окончания к биполярным нейронам. Воздействие света приводит к закрытию натриевых (Na+) каналов, что сопровождается изменением мембранного потенциала фоторецептора, регистрируемого биполярными нейронами. Мри развитии гиперполяризации рецептора высвобождается меньшее количество нейромедиатора, имеющего тормозное действие, а биполярные клетки (и горизонтальные клетки) деполяризуются (возбуждаются). Однако если действие нейромедиатора было бы возбуждающим, происходила бы реполяризация (торможение) данных клеток.
Под действием света происходит гиперполяризация всех палочек, поэтому при высоком уровне освещения их мембранные каналы полностью закрыты, и их вклад в зрение минимален, а зрение обусловлено только функционированием колбочек.
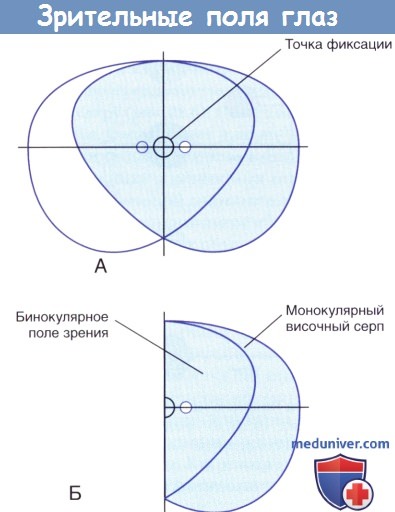
(А) Зрительные поля обоих глаз при фиксации в одной точке. Поле зрения правого глаза окрашено голубым цветом.
(Б) Правое поле зрения. Белая точка обозначает слепое пятно правого глаза.
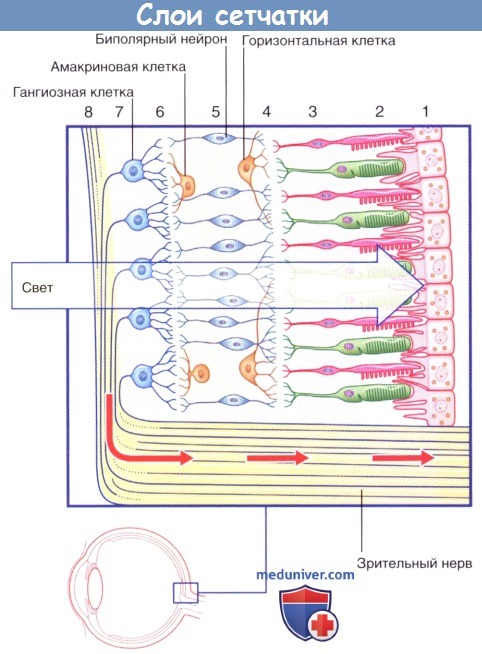
Слои сетчатки:
(1) Пигментный слой. (2) Фоторецепторный слой.
(3) Наружный ядерный слой. (4) Наружный сетчатый слой.
(5) Внутренний ядерный слой. (6) Внутренний сетчатый слой.
(7) Слой ганглиозных клеток. (8) Слой нервных волокон.
2. Палочковые и колбочковые биполярные нейроны:
— Колбочковые биполярные нейроны. Колбочковые биполярные нейроны бывают двух типов. ON-биполярные нейроны возбуждаются (деполяризуются) под действием света и тормозятся нейромедиатором, высвобождаемым в темноте. Они контактируют с ON-ганглиозными клетками. OFF-биполярные клетки реагируют противоположным образом и образуют контакты с OFF-ганглиозными клетками. Как правило, одна колбочка образует синапс с несколькими колбочковыми биполярными нейронами, однако в центральной ямке их отношение составляет 1:1; каждая контактирует только с одной ганглиозной клеткой.
— Палочковые биполярные нейроны. Палочковые биполярные нейроны активируют ON- и OFF-колбочковые ганглиозные клетки косвенно, через амакриновые клетки Один палочковый биполярный нейрон образует синапсы с 15-30 палочками (дополнительные контакты возникают, если реакция распространяется в более центральные отделы).
3. Горизонтальные клетки. Дендриты горизонтальных клеток образуют контакты с фоторецепторами. От периферических ветвей дендритов берут начало аксоноподобные отростки, создающие тормозные контакты с биполярными нейронами.
Функция горизонтальных клеток — торможение биполярных нейронов кнаружи от непосредственной области возбуждения. Возбужденные биполярные клетки и ганглиозные клетки называют «включенными», а заторможенные — «выключенными».
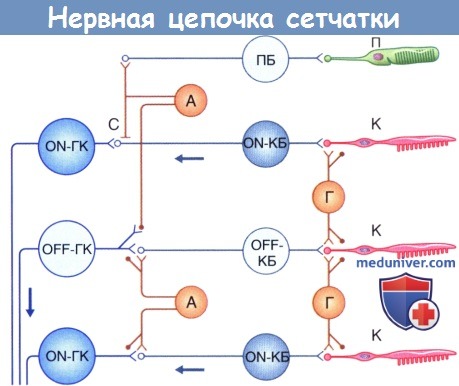
Схема нервной цепочки сетчатки:
А—амакриновая клетка; К—колбочка; КБ—колбочковый биполярный нейрон;
ГК—ганглиозная клетка; Г—горизонтальная клетка; С—соединение (щелевидный контакт);
П—палочка; ПБ—палочковый биполярный нейрон.
4. Амакриновые клетки. Амакриновые клетки не имеют аксонов. Внешне они напоминают осьминога. Все дендриты отходят с одной стороны клетки. Дендритические ветви контактируют с биполярными нейронами и ганглиозными клетками.
Было выделено более десяти различных морфологических типов амакриновых клеток, а также несколько их нейромедиаторов: ацетилхолин, дофамин, серотонин. К возможным функциям этих клеток относят повышение контрастности и регистрацию движений. Амакриновые клетки преобразуют большое количество палочек из OFF в ON в соответствии с типом ганглиозной клетки.
5. Ганглиозные клетки. Ганглиозные клетки образуют синаптические контакты с их биполярными нейронами во внутреннем сетчатом слое. Типичный ответ ганглиозных клеток на возбуждение биполярных нейронов — «от центра к периферии». К центру рецептивного поля относят прямые контакты ганглиозных клеток с фоторецепторами; периферией рецептивного поля считают соединения с прилежащими фоторецепторами через горизонтальные клетки. ON-ганглиозная клетка возбуждается пучком света и тормозится окружающим кольцом света. Торможение осуществляют горизонтальные клетки. OFF-ганглиозная клетка действует по обратному принципу.
— Кодирование цвета. Существует три типа колбочковых фоторецепторов, отличающихся спектральной чувствительностью.
Первый тип фоторецепторов чувствителен к красному цвету (их также называют L-колбочками, так как они регистрируют свет с большей длиной волны — Long), второй тип — к зеленому (М-колбочки), третий—к голубому (их также обозначают как S-колбочки, они составляют приблизительно 5-10 % общего количества колбочек). Чувствительность зависит от строения зрительного пигмента в каждом из типов клеток. Максимальная стимуляция каждого типа колбочек определяет длина волны, однако они отвечают на весьма широкий спектр длин волн, и все три типа колбочек частично дублируют друг друга. Определение цвета зависит не только от типа колбочек, а обусловлено сравнительной активностью различных типов колбочек на определенную длину волны. Группы клеток каждого типа контактируют с ON- или OFF-ганглиозными клетками (обработка цветовой информации начинается в сетчатке и продолжается в латеральном коленчатом ядре и коре полушарий).
Характерная реакция ганглиозных клеток — цветовое противодействие (один цвет возбуждает группу колбочек и их ганглиозную клетку, тогда как «противоположный» цвет тормозит их или их можно рассматривать как взаимно исключающие).
• Ганглиозные клетки, «включенные» для зеленого цвета, «выключены» для красного, а ганглиозные клетки, «включенные» для красного цвета, «выключены» для зеленого.
• Ганглиозные клетки, «включенные» для синего цвета, «выключены» для желтого, ганглиозные клетки, «включенные» для зеленого цвета, «выключены» для желтого.
• Наконец, аналогичный механизм справедлив для черного и белого цветов, а также для яркости изображения.
— Кодирование черного и белого. Белый цвет — это сочетание зеленого, красного и синего. При ярком освещении его кодируют три типа колбочек, взаимодействующих с общей ганглиозной клеткой. ON- и OFF-ганглиозные клетки участвуют в процессах как черно-белого, так и цветового зрения.
В глубоких сумерках, например при свете звезд, активны только палочковые фоторецепторы, и объекты видны в различных оттенках серого. Палочки подчиняются тем же правилам, что и колбочки и обладают центрально-периферическим антагонизмом в отношении белого и черного, а также контактируют как с ON-, так и с OFF-ганглиозными клетками.
Большинство ганглиозных клеток палочек и колбочек — мелкие (Р-клетки — от parvocellular), имеют небольшие рецепторные поля и отвечают за определение формы и цвета. Лишь малая их часть — крупные клетки (М-клетки — от magnocellular), имеют большие рецепторные поля и отвечают за регистрацию движений в поле зрения.
6. Специализация центральной ямки. Относительная плотность колбочек прогрессивно увеличивается, а их размер прогрессивно уменьшается от края центральной ямки к ее центру. Центральная треть центральной ямки (ямочка, foveola) имеет ширину лишь немного более 100 нм и содержит только карликовые колбочки. Для всех колбочек центральной ямки и карликовых колбочек особенно характерны две специфические анатомические особенности, позволяющие передавать максимальное количество информации о форме и цветовых качествах объекта при его внимательном изучении. Во-первых, более поверхностные слои сетчатки отклоняются кнаружи от центра, а их отростки имеют избыточную длину. Это приводит к тому, что наружные две трети ямочки становятся частично перекрытыми телами биполярных клеток, а внутренняя треть ничем не закрыта; свет, отраженный от объекта попадает на колбочки ямочки без какого-либо рассеяния.
Во-вторых, наличие синаптических контактов в отношении 1:1 между карликовыми колбочками и их биполярными нейронами, а также между ними и ганглиозными клетками улучшает точность центральной передачи. Кнаружи от ямочки степень конвергенции «колбочка => биполярная клетка => ганглиозная клетка» прогрессивно увеличивается.
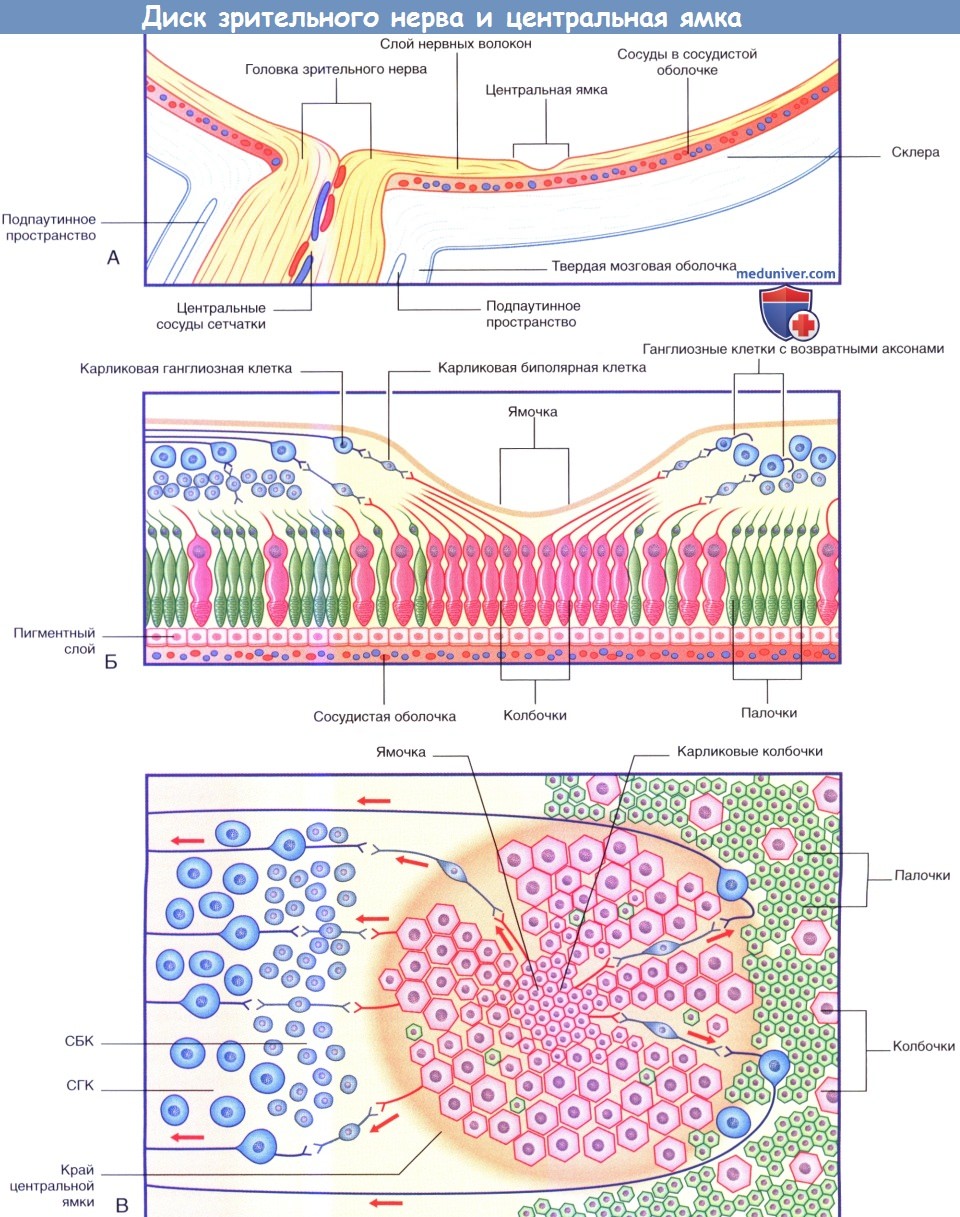
(А) Горизонтальный срез правого глазного яблока на уровне диска зрительного нерва и центральной ямки.
(Б) Увеличенное изображение рисунка А. Возвратные аксоны огибают центральную ямку, как показано на рисунке В.
(В) Поверхность центральной ямки и окружающей сетчатки. Колбочки расположены с интервалами, чтобы показать «цепочечную» последовательность нейронов.
СБК — слой биполярных клеток; СГК — слой ганглиозных клеток.
— Также рекомендуем «Зрительные проводящие пути: зрительный нерв, зрительный путь, коленчато-шпорный путь, первичная зрительная кора»
Редактор: Искандер Милевски. Дата публикации: 21.11.2018
Источник
