
Сетчатки клетки ганглиозные клетки
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг. Аксоны ганглионарных клеток не миелинизированы при прохождении сетчатки, чтобы не препятствовать прохождению света. Далее они покрыты миелиновой оболочкой.
Ганглионарные клетки завершают «трёхнейронную рецепторно-проводящую систему сетчатки»: фоторецептор — биполярный нейрон — ганглионарная клетка.
Функции ганглионарных клеток[править | править код]
Клетки сетчатки связаны между собой сложной сетью возбуждающих, подавляющих и двунаправленных сигнальных связей. Они собирают информацию от всех слоев сетчатки как по вертикальным путям (фоторецепторы — биполяры — ганглионарные клетки), так и по латеральным путям (фоторецепторы — горизонтальные клетки — биполяры — амакриновые клетки — ганглионарные клетки).
Рецептивные поля[править | править код]
На одну ганглионарную клетку приходится от одного до сотни биполярных нейронов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от нескольких до нескольких тысяч фоторецепторов. Каждая ганглионарная клетка суммирует сигналы от большого числа фоторецепторов, что повышает световую чувствительность, но уменьшает разрешение. Фоторецепторы, соединенные с одной ганглионарной клеткой, образуют её рецептивное поле. Рецептивные поля ганглионарных клеток подразделяются на простые и сложные. Простые поля — имеют концентрическую структуру, подобно полям биполярных нейронов. Сложные — разделяются на несколько различных функциональных зон. Рецептивные поля могут перестраиваться, адаптируясь к уровню освещённости и характеристикам световых стимулов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от единиц до десятков тысяч фоторецепторов (палочек и колбочек). В свою очередь, один фоторецептор через биполярные нейроны может быть связан с десятками ганглионарных клеток. В среднем на 100 фоторецепторных клеток приходится одна ганглионарная (т.е., их от 1.2 до 1.5 млн). Чем ближе к центральной ямке глаза, тем меньше фоторецепторов приходится на одну ганглионарную клетку. Поэтому у людей слабое периферическое зрение. В районе центральной ямки, наоборот, высокое разрешение, но менее высокая светочувствительность, поскольку здесь каждый фоторецептор (колбочка) соединен с одной биполярной (карликовой) клеткой, которая в свою очередь соединена лишь с одной ганглионарной.
Типы ганглионарных клеток[править | править код]
Существует всего 18 типов ганглионарных клеток сетчатки.
Большинство относится к трем типам:
1. Парвоганглионарные клетки — карликовые клетки (около 80% от числа всех ганглионарных клеток сетчатки), имеющие средний размер тела и маленькое дерево дендритов, входят в карликовый путь (чувствительный путь, ведущий от глаза к четверохолмию) и связаны с парвоцеллюлярными (мелкоклеточными) слоями латеральных коленчатых тел. С этими клетками связывают высокую остроту зрения и цветовое зрение.
2. Магноклетки — (около 10%) очень разнообразны (малые и большие зонтичные клетки): с большими телами и многочисленными укороченными ветвями, маленькими телами и большим разветвлением дендритов, которые проецируются в крупноклеточные слои латеральных коленчатых тел. Отвечают за востриятие движущихся объектов. Имеют большие рецепторные поля.
3. Кониоцеллюлярные клетки очень мелкие, составляют от 8 до 10% всех ганглионарных клеток сетчатки. Получают сигналы от среднего количества фоторецепторов. Имеют очень большие рецептивные поля. Всегда ON для колбочек синего цвета и OFF для красного и зеленого.
Выделяют ганглионарные клетки, связанные с палочковыми и колбочковыми нейронами, с on- и off-центрами, которые отвечают на световое раздражение деполяризацией или гиперполяризацией соответственно. Дендриты клеток с on-центром разветвляются в подуровне а (пигментном эпителии?), с off-центром в подуровне G (ганглионарном слое?) внутреннего сетчатого слоя. Цветовой канал связан с красным, зеленым и синим типом on/off-ганглионарных клеток. Если красный и зеленый тип ганглионарных on/off-клеток относится к карликовому пути, то синий тип не относится к последнему. On/off-ответы ганглионарных клеток определяются специальными контактами колбочковых биполяров и расположением ганглионарных клеток в соответствующем подуровне внутреннего сетчатого слоя
Светочувствительные ганглионарные клетки[править | править код]
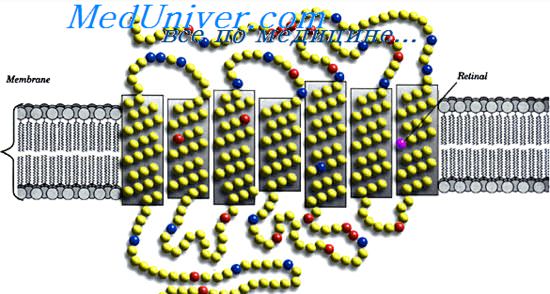
В 1991 году были открыты особые светочувствительные ганглионарные клетки типа ipRGC (intrinsically photosensitive retinal ganglion cells), или mRGC (melanopsin-containing retinal ganglion cells). Они, в отличие от ранее известных ганглионарных клеток, содержат светочувствительный пигмент меланопсин, отличающийся от других фоточувствительных пигментов глаза: родопсина палочек и йодопсина колбочек. И этим они отличаются от других ганглионарных клеток, находящихся в сетчатке глаза, которые не умеют реагировать непосредственно на свет.
Эти светочувствительные ганглионарные клетки — новый, третий тип фоторецепторов сетчатки глаза, помимо известных уже в течение 200 лет палочек и колбочек. Они напрямую возбуждаются под действием света даже при блокировании «классических» фоторецепторов глаза — палочек и колбочек.
Нервные пути от этих ганглиозных (ганглионарных) клеток ведут порождённое в них светом возбуждение от сетчатки к гипоталамусу тремя разными путями, обеспечивая световое управление циркадными ритмами, а также по отдельному нервному пути обеспечивают реакцию сужения зрачка на свет.
Литература[править | править код]
- Ноздрачёв А. Д., Баженов Ю. И., Баранникова И. А., Батуев А. С. и др. Начала физиологии: Учебник для вузов / Под ред. акад. А. Д. Ноздрачёва. СПб.: Лань, 2001. 1088 с.
Ссылки[править | править код]
- Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System
- Photosensitive ganglion cells
- Vision beyond image formation: The role of melanopsin cells in regulating mammalian physiology
- Blind Mice Can «See» Thanks To Special Retinal Cells
- Фоторецепторы и фоторецепция
[1]
[2]
[3]
[4]
[5]
Источник
Амакриновые клетки. Ганглиозные клетки сетчатки
В настоящее время морфологическими или гистохимическими методами идентифицированы около 30 типов амакриновых клеток. Функции некоторых из них охарактеризованы, и все они отличаются друг от друга. Один тип амакриновых клеток является частью прямого пути палочкового зрения, т.е. от палочки к биполярным клеткам, затем к амакриновым и, наконец, к ганглиозным клеткам.
Амакриновые клетки другого типа активно реагируют в начале непрерывного зрительного сигнала, но ответ быстро исчезает. Некоторые амакриновые клетки, наоборот, мощно реагируют на выключение зрительного сигнала, но их ответ также быстро прекращается.
Амакриновые клетки еще одного типа реагируют и на включение, и на выключение света, просто сигнализируя об изменении освещения, независимо от его направления. Есть амакриновые клетки, реагирующие на движение пятна света по сетчатке в определенном направлении; о таких клетках говорят, что они чувствительны к направлению.
В известном смысле большинство амакриновых клеток являются вставочными нейронами, которые помогают анализировать зрительные сигналы, прежде чем они покинут сетчатку.
Ганглиозные клетки
Каждая сетчатка содержит около 100 млн палочек и 3 млн колбочек; однако количество ганглиозных клеток — лишь около 1,6 млн. Таким образом, в среднем 60 палочек и 2 колбочки конвергируют на каждую ганглиозную клетку и волокно зрительного нерва, идущее от ганглиозной клетки к мозгу.
Однако существуют большие различия между периферической и центральной областями сетчатки. По мере приближения к ямке все меньше палочек и колбочек конвергируют на каждое зрительное волокно; кроме того, и палочки, и колбочки становятся тоньше. Эти эффекты постепенно увеличивают остроту зрения в центральной сетчатке. В самом центре — в области центральной ямки — есть только тонкие колбочки (примерно 35000) и совсем нет палочек. К тому же, как показано справа на рисунке, количество волокон зрительного нерва, выходящих из этой части сетчатки, почти равно числу колбочек. Это объясняет высокую степень остроты зрения в центральной сетчатке по сравнению с гораздо меньшей остротой на периферии.
Другим различием между периферией и центром сетчатки является гораздо более высокая чувствительность периферической сетчатки к слабому свету. Отчасти это объясняется тем, что чувствительность палочек в 30-300 раз выше, чем колбочек. Однако эффект значительно усиливается в связи с конвергенцией примерно 200 палочек на одну ганглиозную клетку в периферических областях сетчатки, поэтому сигналы от палочек суммируются, дополнительно усиливая интенсивность стимуляции периферических ганглиозных клеток и исходящих от них волокон зрительных нервов.
Различают три группы ганглиозных клеток, которые обозначают как W-, Х- и Y-клетки. Каждая группа выполняет свою функцию.
Передача палочкового зрения W-клетками. W-клетки составляют примерно 40% общего числа ганглиозных клеток. Они маленькие (диаметром около 10 мкм) и передают сигналы по соответствующим им волокнам зрительного нерва с низкой скоростью, равной около 8 м/сек. Эти ганглиозные клетки возбуждаются в основном от палочек, передающих к ним сигналы через малые биполярные и амакриновые клетки. W-клетки имеют широкие рецептивные поля в сетчатке, т.к. их дендриты широко распространяются во внутреннем слое сетчатки, получая сигналы от обширных областей.
На основании гистологических и физиологических экспериментов показано, что W-клетки, по-видимому, особенно чувствительны к восприятию направленного движения в поле зрения и очень важны для нашего грубого палочкового зрения в условиях темноты.
Передача зрительного образа и цвета Х-клетками. Подавляющее большинство ганглиозных клеток (55%) являются Х-клетками. Они имеют средний диаметр (в пределах от 10 до 15 мкм) и передают сигналы по своим волокнам зрительного нерва со скоростью около 14 м/сек.
Рецептивные поля Х-клеток небольшие, поскольку их дендриты не имеют широкого распространения в сетчатке. В связи с этим сигналы Х-клеток отражают дискретные участки сетчатки. Следовательно, в основном через Х-клетки передаются тонкие детали зрительного образа. Кроме того, поскольку каждая Х-клетка получает сигналы, по крайней мере, от одной колбочки, эти клетки, вероятно, отвечают за все цветовое зрение.
Функция Y-клеток — передача информации о мгновенных изменениях в зрительном образе. Y-клетки — самые большие из всех ганглиозных клеток (диаметром до 35 мкм). Они проводят сигналы к мозгу со скоростью 50 м/сек и выше. Среди ганглиозных клеток они самые малочисленные (около 5% общего количества) и имеют разветвленные дендриты, следовательно, собирают сигналы от обширных областей сетчатки.
Как многие из амакриновых клеток, Y-клетки реагируют на быстрые изменения в зрительном образе (быстрые движения или быстрые изменения освещения), посылая импульсные разряды, длительность которых составляет лишь доли секунды. Эти ганглиозные клетки, вероятно, почти немедленно информируют центральную нервную систему о появлении любого нового зрительного явления в поле зрения, но без высокой точности его локализации, обеспечивая лишь соответствующие сигналы, заставляющие глаза двигаться по направлению к возбуждающему объекту.
— Также рекомендуем «Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке»
Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»:
1. Каскад усиления в сетчатке. Фотохимия цветового зрения
2. Световая и темновая адаптация. Механизмы световой и темновой адаптации
3. Острота зрения. Определение расстояния до объекта глазами
4. Слепота на отдельные цвета. Функция нейронов сетчатки
5. Зрительный путь от колбочек. Нейромедиаторы нейронов сетчатки
6. Функция горизонтальных клеток сетчатки. Возбуждение и торможение биполярных клеток
7. Амакриновые клетки. Ганглиозные клетки сетчатки
8. Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке
9. Зрительные пути. Дорсолатеральное коленчатое ядро таламуса
10. Зрительная кора. Строение первичной зрительной коры
Источник
Оглавление темы «Рецепторный потенциал палочек и колбочек. Рецептивные поля клеток сетчатки. Проводящие пути и центры зрительной системы. Зрительное восприятие.»: М- и Р-типы ганглиозных клеток сетчатки. Магноцеллюлярные (М-клетки) клетки. Парвоцеллюлярные (Р-клетки) ганглиозные клетки сетчатки.Зрительное восприятие происходит в результате согласования друг с другом различных сведений о наблюдаемых объектах. Но на низших иерархических уровнях зрительной системы, начиная с сетчатки глаза, осуществляется независимая переработка информации о форме и глубине объекта, о его цвете и его движении. Паралелльная переработка информации об этих качествах зрительных объектов обеспечивается специализацией ганглиозных клеток сетчатки, которые подразделяются на магноцеллюлярные (М-клетки) и парвоцеллюлярные (Р-клетки).
В большом рецептивном поле относительно крупных М-клеток, состоящем преимущественно из палочек, может проецироваться цельное изображение крупных объектов: М-клетки регистрируют грубые признаки таких объектов и их движение в зрительном поле, отвечая на раздражение всего рецептивного поля непродолжительной импульсной активностью. Клетки Р-типа имеют малые рецептивные поля, состоящие преимущественно из колбочек и предназначенные для восприятия мелких деталей формы объекта или для восприятия цвета. Среди ганглиозных клеток каждого типа имеются как on-нейроны, так и off-нейроны, дающие наиболее сильный ответ на раздражение центра или периферии рецептивного поля. Существование М- и Р-типов ганглиозных клеток позволяет разделить информацию о разных качествах наблюдаемого объекта, которая перерабатывается независимо в параллельных путях зрительной системы: о тонких деталях объекта и о его цвете (пути начинаются от соответствующих рецептивных полей клеток Р-типа) и о движении объектов в зрительном поле (путь от клеток М-типа). — Также рекомендуем «Проводящие пути и центры зрительной системы. Зрительный нерв. Зрительные тракты. Глазодвигательный рефлекс.» |

Источник
Сетчатки ганглиозных клеток ( RGC ) представляет собой тип нейрона , расположенного вблизи внутренней поверхности ( ганглиозных клеток слоя ) из сетчатки в глаза . Он получает визуальную информацию от фоторецепторов с помощью двух промежуточных типов нейронов: биполярные клетки и сетчатки амакринных клеток . Retina амакринные клетки, в частности клетки узкие поля, имеют важное значение для создания функциональных субъединиц в слое ганглиозных клеток и делает его таким образом , что ганглиозные клетки можно наблюдать маленькую точку движущегося на небольшое расстояние. Ганглиозных клеток сетчатки в совокупности передают формирования изображения и не-формирования изображения визуальную информацию от сетчатки в форме потенциала действия в некоторых регионах в таламус , гипоталамус и мезенцефалона или среднего мозга .
Сетчатке ганглиозных клеток существенно различаются с точки зрения их размера, соединений, и ответы на визуальной стимуляции , но все они имеют определяющее свойство иметь длинный аксон , который проходит в мозг. Эти аксоны образуют зрительный нерв , зрительный перекрест , и оптический тракт .
Небольшой процент ганглиозных клеток сетчатки не способствует мало или вообще ничего видению, но сами по себе светочувствительным; их аксоны образуют retinohypothalamic тракта и способствуют циркадных ритмов и зрачкового светового рефлекса , изменения размера зрачка.
Шесть типов нейронов сетчатки являются биполярные клетки , ганглиозные клетки, горизонтальные клетки , сетчатка амакринные клетки , и стержень и конус фоторецепторов .
функция
Есть около 0,7 до 1500000 ганглиозных клеток сетчатки в сетчатке глаза человека. С около 4,6 млн колбочек и 92 млн палочек , или 96,6 миллионов фоторецепторов в сетчатке, в среднем каждый ганглиозных клеток сетчатки принимает входные сигналы от около 100 палочек и колбочек. Тем не менее, эти цифры сильно различаются между отдельными лицами и в зависимости от расположения сетчатки глаза. В ямке ( в центре сетчатки), один ганглиозных клеток будет взаимодействовать с всего лишь пять фоторецепторов. В крайней периферии (концы сетчатки), один ганглиозные клетки будут получать информацию из многих тысяч фоторецепторов.
Ганглиозные клетки сетчатки самопроизвольно срабатывали потенциалы действия на базовую скорости в то время как в состоянии покоя. Возбуждение сетчатки приводит ганглиозные клетки к увеличению скорости обжига , а результаты ингибирования в депрессивной скорости стрельбы.
Ложный-цветное изображение плоской монтажа сетчатки крыс наблюдает через флуоресцентный микроскоп при 50-кратном увеличении. Зрительный нерв был введен с флуорофором, в результате чего ганглиозных клеток сетчатки флуоресцировать.
Типы
Три группы
- W-ganglion- небольшого, 40% от общего количества, широких полей в сетчатке, возбуждения от стержней, обнаружение движения направления в любой точке области.
- Х-ganglion- средний диаметр, 55% от общего числа, небольшое поле, цветовое зрение. Устойчивый ответ.
- Y- ганглий cells- по величине, 5%, очень широкое поле дендритного, реагирует на быстрое движение глаз или быстрое изменение интенсивности света. Переходный ответ.
На основе их проекций и функций, существует, по крайней мере, пять основных классов ганглиозных клеток сетчатки:
- Малыш клетки (Parvocellular, или Р путь; P клетка )
- Зонтик клетки (магноцеллюлярная, или М пути; М — клетка )
- Bistratified клетки (Koniocellular, или К пути)
- Фоточувствительные клетки ганглия
- Другие клетки ганглия , выступающие в двухолмия для движений глаз ( саккады )
карлик
Midget ганглиозных клеток сетчатки проект к parvocellular слоев в латеральном ядре коленчатого . Эти клетки известны как карликовые ганглиозные клетки сетчатки, на основе небольших размеров их дендритных дерев и клеточные тел. Около 80% всех ганглиозных клеток сетчатки являются карликовые клетки в parvocellular пути . Они получают сигналы от относительно небольшого числа палочек и колбочек. Они имеют медленную скорость проводимости , и реагировать на изменения в цвете , но реагируют слабо на изменения в контрасте , если изменение не велико (Кандель и др., 2000). Они имеют простой центральные тыловые рецептивные поля , где центр может быть либо ON или OFF , а обрамление наоборот.
зонтик от солнца
Зонтик ганглиозных клеток сетчатки проект к магноцеллюлярным слоям бокового коленчатого тела. Эти клетки известны как зонтик ганглиозных клеток сетчатки, основанный на крупных размерах их дендритные деревьев и клеточных тел. Около 10% всех ганглиозных клеток сетчатки являются пляжный зонт клетки, и эти клетки являются частью магноцеллюлярного пути. Они получают сигналы от относительно многих палочек и колбочек. Они имеют скорость быстрой проводимости, и может реагировать на малоконтрастных раздражители, но не очень чувствительны к изменениям в цвете (Кандель и др., 2000). Они имеют гораздо большие рецептивные поля , которые тем не менее являются также центром объемного звучания.
Bistratified
Bistratified ганглиозных клеток сетчатки проект к koniocellular слоев бокового коленчатого тела. Bistratified ганглиозные клетки сетчатки были выявлены сравнительно недавно. Koniocellular означает «клетки , как малые , как пыль»; их небольшой размер делает их трудно найти. Около 10% всех ганглиозных клеток сетчатки являются bistratified клетки, и эти клетки проходят через koniocellular пути. Они получают сигналы от промежуточных чисел палочек и колбочек. Они имеют умеренное пространственное разрешение, скорость умеренной проводимости, и может реагировать на умеренно-контрастные раздражители. Они могут быть вовлечены в цветовом зрении. Они имеют очень большие рецептивные поля , которые имеют только центры (не окружает) и всегда находятся в голубой конус и выключенный как красный и зеленый конус.
Фоточувствительные клетки ганглия
Светочувствительные ганглиозные клетки , в том числе , но не ограничиваясь гигантскими ганглиозными клетки сетчатки, содержат свой собственный фотопигмент , меланопсин , что делает их непосредственно реагируют на свет , даже в отсутствии палочек и колбочек. Они проецируют, среди других областей, супрахиазматическое ядро (SCN) через retinohypothalamic тракта для установления и поддержания циркадных ритмов . Другие ганглиозные клетки сетчатки , выступающие в латеральное ядро коленчатого (LGN) включают в себя клетку делает связь с ядром Эдингер-Вестфаль (EW), для контроля светового рефлекса зрачка и гигантских ганглиозных клеток сетчатки .
физиология
Большинство зрелых ганглиозных клеток способны стрелять потенциалы действия на высокой частоте из — за их экспрессии К v 3 калиевых каналов.
биология развития
Ниже приведен очень краткий обзор. Для эталонного источника, обратитесь к разделу «Подключение сетчатки в мозг» по Эрскин и др. и «сетчатка Axon рост на хиазмах» Петрос TJ и др. Большая часть информации, выделена ниже происходит от этих двух прекрасных обзоров.
Сетчатка рост: начало
Сетчатку клетки нервного узла (РГК) родились между 11 -й день эмбрионального (Е11) и пост-natals (PN) нулевой день (Pn0) в мыши и между неделю 5 и 18 недели в утробе матери в развитии человека. У млекопитающих, ГКС , как правило , добавляет в начале в дорсальном-центральном аспекте глазного бокала , который расположен в центре глаза. Затем рост RC будет подметать вентрально и периферические оттуда, в волнообразном рисунке. Этот процесс зависит от множества факторов, начиная от сигнальных факторов , таких как FGF3 и FGF8 для надлежащего ингибирования сигнального пути Notch. Самое главное, что bHLH (основная спираль-петля-спираль) -области , содержащий фактор транскрипции Atoh7 и ее вниз по течению эффекторов, такие как Brn3b и Isl-1 , работа по содействию RGC выживания и дифференциации . «Дифференцировка волна» , что приводит в развитие ГКС через сетчатку также регулируется , в частности, bHLH факторы Neurog2 и Ascl1 и FGF / сигнализации Shh, вытекающих из периферии.
Рост в пределах сетчатки ганглиозных клеток (оптическое волокно) слой
Ранние ГКС — предшественники, как правило , распространяются процессы с подключением к внутренним и внешним лимитирующих мембран сетчатки с наружным слоем , прилегающей к пигментного эпителия сетчатки (RPE) и внутренней прилегающей к будущему стекловидного тела. Клеток сомы будет тянуть к ПЭС, подвергаются терминальной дифференцировки, а затем мигрируют в обратном направлении внутренней ограничивающей мембраны. Он будет распространяться аксоном в сетчатке слоя ганглиозных клеток. Этот процесс , вероятно , опосредуются щелевой-Robo сигнализация, который управляет втягиванием апикальной процессе RGC.
ГКС будет расти вдоль глиальных конечных ног , расположенных на внутренней поверхности стекловидного (сторона ближе к будущему стекловидного тела). Нейронные молекулы клеточной адгезии (N-CAM) будет посредник этого вложения через гомофильный (N-САМ связывается только с этим изотопом N-CAM: А с А, В с В, никогда А с В) взаимодействиях. Сигнализации щелевые также играет определенную роль, предотвращая РКИ от растущей на слои за пределами оптического волокнистого слоя.
Аксоны образуют РКИ будут расти и распространяться в направлении диска зрительного нерва , где они выходят из глаз. После того, как дифференцировано, они граничат с ингибирующей периферийной областью и центральной областью привлекательной, способствуя тем самым расширение аксона в направлении диска зрительного нерва. CSPGs существует вдоль нейроэпителии сетчатки (поверхностей , по которой лежит ГКС) в периферическом высоком центральном низком градиенте. Прорезь также выражается в аналогичной схеме, секретируемый из клеток в линзе. Молекулы адгезии, такие как N-CAM и L1, будут способствовать росту централизованно , а также помогут правильно расположенному пучок (расслоение) аксонам RGC вместе. Тсс выражаются в высоком, низком центральном периферическом градиенте, способствуя центрально-проецированием расширения аксонов ГКСА через исправленные-1, основной рецептор для Shh, опосредованной сигнализация.
Рост в и через зрительный нерв
ГКС выход из сетчатки слоя ганглиозных клеток через диск зрительного нерва, который требует 45 ° поворота. Это требует сложных взаимодействий с оптическими дисками глиальных клетками, которые экспрессируют будут локальные градиенты Netrin-1, морфоген, который будет взаимодействовать с удаленным в колоректальном раке (DCC) рецептором на конусах роста аксонов RGC. Это морфоген изначально привлекает РКИ аксоны, но затем, через внутреннее изменение в конусе роста RGC, Netin-1 становится отталкивающим, толкая аксон от диска зрительного нерва. Это опосредовано через зависимый механизм цАМФа. Кроме того, CSPGs и сигнализация Эф / эфрин также могут быть вовлечены.
ГКС будет расти вместе конечной глиальные клетки ноги в зрительном нерве. Эти глии секретируют отталкивающее Semaphorin 5ы и щель в объемной моде, охватывающая зрительный нерв , который гарантирует , что они остаются в зрительном нерве. Vax1 , фактор транскрипции , выражается вентральных диэнцефалона и глиальных клеток в области , где формируется перекрест, и он также может секретироваться контролировать образование перекреста.
Рост при зрительных нервах
Когда РКИ подойти зрительный перекрест, точка, в которой два зрительных нервах встретиться, на вентральный диэнцефалоне вокруг E10-E11 у мышей, они должны принять решение пересекающегося под прямой углом к контралатеральному зрительному тракту или остаются в ипсилатеральном зрительном тракте. У мышей, около 5% от ГКС, в основном тех, кто прибывает из вентральной-временной серп (VTC) области сетчатки, останется ипсилатеральная, а остальные 95% ГКС будет пересекать. Это в значительной степени контролируется степенью бинокулярного перекрытия между двумя полями зрения в обоих глазах. Мышь не имеет значительное перекрытие, в то время как люди, которые делают, будут иметь около 50% от пересекаться РКИ и 50% останутся ипсилатеральной.
Построение отталкивающего контур перекреста
После того, как ГКС достигает перекрест, глиальные клетки, поддерживающие их будут меняться от intrafascicular к радиальной морфологии. Группа диэнцефальных клеток, которые экспрессируют поверхностный антиген клеточной стадии-эмбриональный антиген (ССЭА) -1 и CD44 образует перевернутую V-образную форму. Они установят заднюю сторону границы хиазмов. Кроме того, передача сигналов щелевая важно здесь: Гепарин сульфат протеогликаны (HSPGs), белки ЕСМ, будет якорь морфоген щелевой в определенных точках на границе задней перекреста. ГКС начнет выражать Robo, рецептор для слейте, в этой точке, таким образом, облегчая отталкивание.
В противоположных выступающих РКАХ
RGC аксона, путешествующая в контралатеральном зрительном тракт должна пересекаться. Тсс играет роль в этом. Это выражается вдоль средней линии в брюшном диэнцефалоне, обеспечивая отталкивающий кий, чтобы предотвратить РКИ от пересечения средней линии эктопический. Однако, дыра образуется в этом градиенте, таким образом, позволяя ГКС в расположенном крестообразно (исследование все еще активно сделано, чтобы понять механизм позади этого изолированной облитерации).
Молекулы, опосредующая привлекательность включают в себя NrCAM, которая выражается растущим ГКСОМ и средней линии глии и действует вместе с Sema6D, опосредованной через рецептор плексин-A1. VEGF-A освобождается от средней линии направляет РГК принять контралатеральной путь, опосредованный рецептором Neuropilin-1 (NRP1). цАМФ, кажется, очень важную роль в регуляции производства белка NRP1, регулируя тем самым роста ответ конусы к VEGF-A градиента в перекреста.
Ипсилатеральное проектирование РОК
Единственный компонент , у мышей , выступающая ипсилатерально является ГКСОМ из вентрального-временного серпа в сетчатке, и только потому , что они выражают Zic2 фактор транскрипции. Zic2 будет способствовать экспрессии рецептора EphB1 тирозинкиназы, которые посредством прямой передачи сигналов (см обзор, Xu и др.) Будут связываться с эфрин В2 лиганда , выраженной средней линии глии и отталкиваться , чтобы повернуть в сторону от перекреста. Некоторый ГКС VTC будет выступать контралатерально , поскольку они выражают фактор транскрипции Островок-2 , который является негативным регулятором Zic2 производства.
Тсс играет ключевую роль в поддержании РОК аксоны ипсилатеральный, а также. Тсс выражается контралатерально выступающими ГКС и средней линии глиальных клеток. Вос или брат CDO (САМ, связанный с / подавляются онкогенами), со-рецептор для Shh, который влияет на Shh сигнализации через pTCH1, кажется, опосредует это отталкивание, как это только на конусах роста, поступающих из ипсилатерально выступающего ГКСА.
Другие факторы, влияющие на рост ипсилатерального RGC включают семейство Teneurin, которые являются трансмембранными белками адгезии, которые используют гомофильные взаимодействия для контроля ориентации, и Nogo, которая выражается с помощью средней линии радиальной глии. Рецептора Nogo экспрессируется только VTC ГКС. Роль Nogo в передаче сигналов на перекресте, однако, все еще в стадии активного исследования.
Наконец, другие факторы транскрипции, кажется, играют важную роль в изменении. Например, Foxg1, называемые также Мозг-фактором 1, и Foxd1, называемый также Мозг Фактор 2, являются крылатой-спираль факторы транскрипции, которые экспрессируются в носовом и временных оптических чашках и оптические везикулы начинают с выпячиваться из нервной трубки. Эти факторы также экспрессируется в вентральном диэнцефалоне с Foxd1 выражается вблизи перекреста, в то время как Foxg1 выражаются более ростральным. Они по-видимому, играют важную роль в определении ипсилатеральной проекции, изменяя экспрессию рецептора Zic2 производства и EphB1.
Рост в зрительном тракте
После того, как из зрительных нервов, ГКС будет распространяться dorsocaudally вдоль вентральной поверхности диэнцефального делая зрительный тракт, который будет вести их к двухолмия и латерального ядра коленчатом у млекопитающих, или тектума в низших позвоночных. Sema3d , кажется, способствуют росту, по крайней мере , в ближнем зрительном тракте и цитоскелета повторные механизмы на уровне конуса роста является значительным.
миелинизация
У большинства млекопитающих, аксоны ганглиозных клеток сетчатки не миелиновые , где они проходят через сетчатку. Однако, части аксонов , которые находятся за пределами сетчатки, являются миелинизированными. Эта модель миелинизации функционально объясняется относительно высокой непрозрачностью миелина миелиновые аксонов , проходящих через сетчатку будет поглощать часть света , прежде чем он достигнет фоторецепторов слоя, снижая качество зрения. Есть заболевание глаз человека , где это, на самом деле, происходит. В некоторых позвоночных животных, например , курица, ганглиозные клетки аксона являются миелиновой внутри сетчатки.
Смотрите также
- ганглиозные клетки
- Восприимчивость поле
Рекомендации
внешняя ссылка
- Диаграмма на mit.edu
- Обзор и диаграммы на webexhibits.org
- Neuronbank Wiki страница на ГКС
- NIF Поиск — сетчатка ганглиозных клеток через информационные рамки Neuroscience
Источник

