
Превращение энергии в сетчатке
К
 вант
вант
света
Ф
 оторецепторы
оторецепторы
сетчатки
Активация
родопсина: свет, поглощаемый родопсином,
инициирует ответ в каскаде цГМФ

Каталитическая
активация G-белка
родопсином

Активация
циклической фосфодиэстеразы

Г
 идролиз
идролиз
цГМФ активированной цГМФ-фосфодиэстеразой
З
 акрытие
акрытие
цГМФ зависимыхNa+
каналов
Г
 иперполяризация
иперполяризация
фоторецепторов сетчатки
Передача
сигнала на биполярные клетки: изменение
характера секреции медиатора в синапсе
между внутренним сегментом рецепторной
клетки и дендритом биполярной клетки
Механизм
восстановленияисходного
темного состояния фоторецептора связан
с повышением концентрации цГМФ (в
темновую фазу с участием алкагольдегидрогеназы
+ НАДФ). Снижение проницаемости натриевых
каналов при действии света сопровождается
также снижением поступления в клетку
ионов Са2+,
уменьшение
его концентрации в клетке ведет к
повышению активности фермента
гуанилатциклазы, увеличивающей скорость
образования цГМФ из ГМФ. Уменьшение
концентрации ионов Са2+
также
приводит к снижению активности фермента
фосфодиэстеразы, разрушающей цГМФ. В
результате всего этого концентрация
цГМФ внутри клетки повышается и цГМФ
вновь связывается с ионным каналом
плазматической мембраны, открывая его.
Через открытый канал внутрь клетки
вновь начинают входить Na+
и Ca2+,деполяризуя
мембрану рецептора и переводя его в
«темновое» состояние. Из пресинаптического
окончания деполяризованного рецептора
вновь ускоряется выход медиатора.
Предполагается, что от уровня кальция
существенно зависит скорость восстановления
чувствительности(адаптации) фоторецепторов
как в условиях высокой , так и низкой
освещенности.
Т.о.,
поглощение света, молекулами фотопигмента
приводит к снижению проницаемости для
Nа+,
что сопровождается гиперполяризацией,
т.е. возникновением рецепторного
потенциала. Гиперполяризационный
рецепторный потенциал, возникший на
мембране наружного сегмента,
распространяется затем вдоль клетки
до ее пресинаптического окончания и
приводит к уменьшению скорости выделения
медиатора —глутамата.
Кроме глутамата нейроны сетчатки могут
синтезировать и другие нейромедиаторы,
такие как ацетилхолин,
дофамин, глицин и ГАМК.
На уровне рецепторов происходит
торможение, и сигнал колбочки перестает
отражать число поглощенных фотонов, а
несет информацию о распределении и
интенсивности света, падающего на
сетчатку в окрестностях рецептора.
Механизмы
восприятия и преобразования световых
сигналов колбочками схожи с таковыми
для палочек. Основные отличия реагирования
колбочек на световые воздействия связаны
с их более низкой (~в 70 раз) фоточувствительностью
по сравнению с палочками.
Фоторецепторы
связаны между собой — электрическими
(щелевыми) контактами. Эта связь
избирательная: палочки связаны с
палочками, а колбочки с колбочками.
Эти
ответы от фоторецепторов сходятся на
горизонтальные клетки, которые приводят
к деполяризации в соседних колбочках
возникает отрицательная обратная связь,
которая повышает световой контраст.
Существует
три типа нейронов сетчатки — биполярные,
горизонтальные и амакриновые клетки.
Биполярные
клетки непосредственно связывают
фоторецепторы с ганглиозными клетками,
т.е. осуществляют передачу информации
через сетчатку в вертикальном направлении.
Горизонтальные и амакриновые клетки
передают информацию по горизонтали
(рис. 25.).
Биполярные
клетки
занимают в сетчатке стратегическую
позицию, поскольку все сигналы, возникающие
в рецепторах поступающие к ганглиозным
клеткам, должны пройти через них.
Экспериментально было доказано, что
биполярные
клетки имеют рецептивные поля
в которых выделяют центр
и периферию
(Джон Даулинг и др., Гарвардская медицинская
школа).
Рецептивное
поле — совокупность рецепторов, посылающих
данному нейрону сигналы через один или
большее число синапсов.
Размер
рецептивных полей: d=10
мкм
или 0,01 мм — вне центральной ямки.
 Р
Р ис.
ис.
25. Синаптические связи в сетчатке (схема
по Е.Бойкоту, Дж. Даулингу): 1 — пигментный
слой;
2
— палочки; 3 — колбочки; 4 — зона расположения
наружной погранич-ной мембраны; 5 —
горизонтальные клетки; 6 — биполярные
клетки; 7 — амакриновые клетки; 8 — глия
(мюллерово
волокно); 9 – ганг-лиозные клетки; 10 —
зона расположения внутренней пограничной
мембраны; 11 – синапсы между фоторецеп-торами,
биполярными и горизонтальными нейронами
в наружном сетчатом слое; 12 — синапсы
между биполярными, амакриновыми и
ганлиозными клетками во внутреннем
сетчатом слое.
В
самой ямке d=2,5мкм
(благодаря
этому мы способны различать две точки
при видимом расстоянии между ними лишь
0,5 угловых минут-2,5мкм — если сравнить,
то это монета в 5 копеек на расстоянии
около 150 метров).
Начиная
с уровня биполярных клеток нейроны
зрительной системы дифференцируются
на две группы (рис. 26), противоположным
образом реагирующие на освещение и
затемнение:
1 —
клетки, возбуждающиеся при освещении
и тормозящиеся при затемнении —
«on»-нейроны
и
–
клетки,
возбуждающиеся
при затемнении и тормозящиеся при
освещении
— «off»-нейроны.
Клетка с on-центром
разряжается с заметно повышенной
частотой. Если слушать разряды такой
клетки через громкоговоритель, то
сначала вы услышите спонтанную
импульсацию, отдельные случайные щелчки,
а затем после включения света, возникает
залп импульсов, напоминающий пулеметную
очередь.
Наоборот,
в клетках с off-реакцией (при выключении
света — залп импульсов). Такое разделение
сохраняется на всех уровнях зрительной
системы, до коры включительно.
Р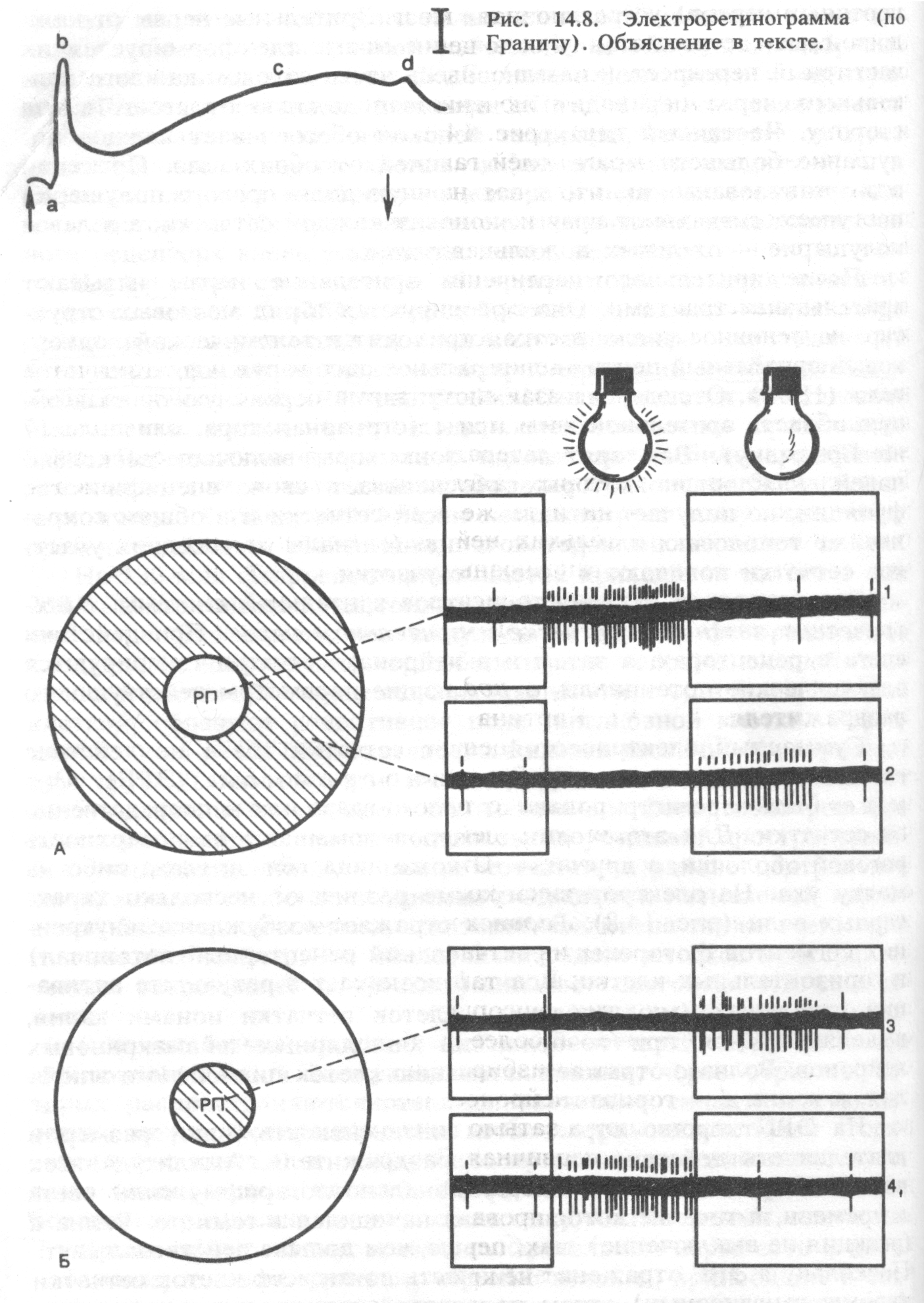 ис.
ис.
26. Концентрические рецептивные поля
(РП) двух ганглиозных клеток.
Тормозные
зоны рецептивных полей заштрихованы.
Показаны реакции на включение (1 и 4)
и выключение(2 и 3) света при
стимуляции световым пятном центра РП
(1 и 3) и его периферии (2 и 4).
А
— «on»-нейроны
Б
— «
off»-нейроны
В
пределах самой сетчатки передача
информации осуществляется безимпульсным
путем
(распространением и транссинаптической
передачей градуальных потенциалов).
В
горизонтальных, биполярных и амакриновых
клетках переработка сигнала происходит
путем медленных изменений мембраны
потенциалов (тонический ответ). ПД не
генерируется.
Ответы
палочек, колбочек и горизонтальных
клеток являются гиперполяризующими, а
ответы биполярных клеток могут быть
как гиперполяризующие, так и деполяризующие.
Амакриновые клетки создают деполяризующие
потенциалы.
Чтобы
понять, почему это так, следует представить
себе влияние малого светлого пятна.
Рецепторы активны в темноте, а свет,
вызывая гиперполяризацию, уменьшает
их активность. Если синапс
возбуждающий,
биполяр
будет активироваться в темноте,
а инактивироваться
на свету;
если же синапс
тормозной, биполяр в темноте тормозится,
а на свету, выключая рецептор, снимает
это торможение, т.е. биполярная клетка
активируется. Таким
образом, является ли рецепторно-биполярный
синапс возбуждающим или тормозным,
зависит от выделяемого рецептором
медиатора.
В
передаче сигналов от биполярных клеток
на ганглиозные участвуют горизонтальные
клетки, которые передают информацию от
фоторецепторов к биполярным клеткам и
далее к ганглиозным.
Горизонтальные
клетки
отвечают на свет гиперполяризацией с
ярко выраженной пространственной
суммацией. Горизонтальные клетки не
генерируют нервных импульсов, но мембрана
обладает нелинейными свойствами,
обеспечивающими безимпульсное проведение
сигнала без затухания.
Клетки
делятся на два типа: В и С. Клетки В-типа,
или яркостные, всегда отвечают
гиперполяризацией вне зависимости от
длины волны света. Клетки С-типа, или
хроматические делятся на двух- и
трехфазные. Хроматические клетки
отвечают или гипер-, или деполяризацией
в зависимости от длины стимулирующего
света.
Двухфазные
клетки бывают либо красно-зеленые
(деполяризуются красным светом,
гиперполяризуются зеленым), либо
зелено-синие (деполяризуются зеленым
светом, гиперполяризуются синим).
Трехфазные клетки деполяризуются
зеленым светом, а синий и красный свет
вызывает гиперполяризацию мембраны.
Амакриновые
клетки,
регулируют синаптическую передачу на
следующем этапе от биполяров к ганглиозным
клеткам. Дендриты амакриновых клеток
разветвляются во внутреннем слое, где
контактируют с отростками биполяров и
дендритами ганглиозных клеток. На
амакриновых клетках оканчиваются
центробежные волокна, идущие из головного
мозга.
Амакриновые
клетки генерируют градуальные и
импульсные потенциалы (фазный характер
ответа). Эти клетки отвечают быстротекущей
деполяризацией на включение и выключение
света и демонстрируют слабый
пространственный антагонизм между
центром и периферией.
Выходными
элементами сетчатки являются ганглиозные
клетки.
В
сетчатке на 110
— 123 млн
палочек и 6
— 7 млн колбочек
приходится около 1,6 млн ганглиозных
клеток. В среднем на одну ганглиозную
клетку конвергирует 60
палочек и 2 колбочки.
Ганглиозная
клетка сетчатки – это первый нейрон
«классического» типа в прямой цепи
передачи информации от фоторецепторов
к мозгу.
Возбуждение
ганглиозных клеток сетчатки приводит
к тому, что по их аксонам (волокнам
зрительного нерва) в мозг устремляются
нервные импульсы.
Различают
3 типа ганглиозных клеток.
Среди ганглиозных клеток выделяют:
Тонические
нейроны (Х–нейроны
— 55%)
которые
возбуждаются адекватно световыми
стимулами и отвечают в течении всего
времени действия стимула.Фазические
(Y–нейроны
— 5%)
самые крупные ганглиозные клетки
кратковременный
ответ длиной в несколько секунд,
реагируют на движущиеся стимулы.
III.
W–нейроны
(составляют 40%
от общего числа)
Получают сигналы от палочек и отвечают
за сумеречное зрение.
В
рецептивном поле ганглиозных клеток
выделяют центр и переферию (оn-реакция),
(off-реакция),
(on—off-реакции).
Т.е.
клетки (рис.26.) противоположным образом
реагирующие на освещение и затемнение:
1
— клетки, возбуждающиеся при освещении
и тормозящиеся при затемнении «on»-нейроны
2
— клетки возбуждающиеся при затемнении
и тормозящиеся при освещении — off-нейроны.
Антогонистическая
организация РП ганглиозных полей
обусловлена наличием 2-х
путей проведения в сетчатке.
Центр
обладает максимальной световой
чувствительностью, а на краю чувствительность
снижается.
Реакции
на возбуждение центра связана с
вертикальным путем проведения от
фоторецептора к биполярным клеткам и
далее к ганглиозным.
Реакции
на раздражение периферии обусловлены
горизонтальным путем проведения.
Конвергенция
и дивергенция связей в сетчатке составляют
основу рецептивных полей ганглиозных
клеток сетчатки.
Источник
Задняя стенка глаза выстлана эпителием, содержащим черный пигмент меланин, который препятствует отражению света и поглощает его излишки. К пигментному эпителию примыкает слой фоторецепторов, а перед ним расположены слои горизонтальных, биполярных, амакриновых и ганглиозных клеток, чьи аксоны образуют зрительный нерв. В связи с инверсией сетчатки решаются две задачи: 1) обеспечение максимальной прозрачности клеток, расположенных на пути прохождения света к фоторецепторам, 2) выделение в слое фоторецепторов промежутка для прохождения волокон зрительного нерва.
Решение первой задачи заключается в анатомическом устройстве центральной ямки, где биполярные и ганглиозные клетки сетчатки располагаются вокруг нее, а их относительно прозрачные безмиелиновые волокна не мешают прохождению света к фоторецепторам. В других областях сетчатки световые лучи отчасти рассеиваются при прохождении сквозь тела клеток, расположенных перед фоторецепторами, но в центральной ямке кванты света без помех попадают на фоторецепторы, представленные здесь преимущественно колбочками. При направленном на определенный объект зрительного поля внимании его изображение всегда фокусируется в центральной ямке для обеспечения максимальной остроты зрения.
Сосок зрительного нерва образован аксонами ганглиозных клеток, отсутствие в этой области фоторецепторов делает ее не чувствительной к свету, в связи с чем она получила название слепого пятна. На соски правого и левого глаза благодаря диспарации попадают неодинаковые части проецируемого изображения, поэтому в условиях естественного зрительного восприятия пробел изображения на сетчатке не осознается. Кроме того, быстрые движения глаз позволяют получать нужную информацию о любой области рассматриваемого изображения, и отсутствие сигналов от слепого пятна замещается информацией, поступившей от ближайших к нему фоторецепторов. Скотопическая и фотопическая системы сетчатки
Фоторецепторный слой сетчатки человека образован примерно 130 миллионами клеток, из которых около семи миллионов являются колбочками, основная масса которых сосредоточена в области центральной ямки, а все остальные фоторецепторы представлены палочками. У обеих разновидностей фоторецепторов существуют три функциональные области: 1) наружный, или внешний, сегмент, ориентированный в направлении эпителиального пигментного слоя и содержащий зрительный пигмент; 2) внутренний сегмент, в котором расположено клеточное ядро и происходят биохимические процессы, связанные с жизнедеятельностью клетки; 3) синаптические окончания, предназначенные для передачи информации от фоторецепторов к биполярным клеткам с помощью медиатора глутамата.
Зрительный пигмент палочек родопсин состоит из двух компонентов: это молекула ретиналя, образующаяся из витамина А и способная поглощать свет, а также крупная белковая молекула опсина, не поглощающая свет. Молекула опсина представляет собой извитую цепь из 348 аминокислот, которая семь раз проходит через мембрану зрительного диска, образованного из клеточной мембраны фоторецептора. В наружном сегменте фоторецептора имеется большое количество таких дисков, расположенных подобно стопке поставленных друг на друга монет. Ретиналь существует в темноте как 11-цис-ретиналь, такая форма изомера идеально соответствует упорядоченному расположению аминокислот в опсине. Энергия поглощенных фотонов превращает ретиналь в 11-транс-изомер, что приводит к конформационным изменениям молекулы опсина и превращению родопсина в нестабильный метародопсин, который сразу же распадается на ретиналь и опсин. Таким образом, действие света уменьшает концентрацию родопсина в фоторецепторе, что приводит к изменениям активности вторичных посредников и величины мембранного потенциала фоторецептора. В темноте происходит ферментативный ресинтез расщепленного родопсина, для которого используется витамин А, поступающий в организм человека с пищей.
Родопсин наиболее чувствителен к электромагнитным волнам длиной около 500 нм, но хорошо поглощает и другие волны в диапазоне от 400 до 600 нм. Способность родопсина поглощать волны почти всего светового диапазона позволяет палочкам обеспечить только ахроматическое, т. е. черно-белое, зрение и лишает их возможности различать цвет. Высокое содержание зрительного пигмента в палочках и его способность суммировать фотоны, поглощенные в течение около 100 мс, делает палочки наиболее чувствительными фоторецепторами сетчатки. При слабом сумеречном освещении зрение обеспечивают только палочки, способные возбуждаться вследствие поглощения всего лишь одного фотона. Палочки образуют ско- топическую систему, или систему ночного зрения.
Зрительные пигменты колбочек подобны родопсину палочек и состоят

Рис. 17.7. Спектры поглощения четырех разновидностей фоторецепторов.
Зрительный пигмент палочек родопсин (Р) имеет максимум поглощения световых волн длиною 496 нм, но способен также к поглощению коротких и длинных волн светового диапазона. Зрительный пигмент колбочек, чувствительных к синему цвету (С), имеет максимум поглощения 419 нм и не поглощает длинные волны оптического диапазона. Пигмент колбочек, чувствительных к зеленому цвету (3), имеет максимум поглощения при 531 нм, а пигмент чувствительных к красному цвету колбочек (К) максимально поглощает волны длиной 596 нм.
из светопоглощающей молекулы ретиналя и опсина, который отличается составом аминокислот от белковой части родопсина. Кроме того, колбочки содержат меньшее, чем палочки, количество зрительного пигмента, и для их возбуждения требуется энергия нескольких сотен фотонов. Поэтому колбочки активируются лишь при дневном или достаточно ярком искусственном освещении, они образуют фотопическую систему, или систему дневного зрения.
В сетчатке человека существуют три типа колбочек, различающихся между собой по составу аминокислот в опсине зрительного пигмента. Различия в белковой части молекулы определяют особенности взаимодействия каждой из трех форм опсина с ретиналем и специфическую чувствительность к световым волнам разной длины (рис. 17.7). Колбочки одного из трех типов максимально поглощают короткие световые волны с длиной 419 нм, что необходимо для восприятия синего цвета. Другой тип зрительного пигмента наиболее чувствителен к волнам средней длины и имеет максимум поглощения при 531 нм, он служит для восприятия зеленого цвета. Третий тип зрительного пигмента максимально поглощает длинные волны с максимумом при 559 нм, что позволяет воспринимать красный цвет. Наличие трех типов колбочек обеспечивает человеку восприятие всей цветовой палитры, в которой существует свыше семи миллионов цветовых градаций, тогда как скотопическая система палочек позволяет различать лишь около пятисот черно-белых градаций.
Рецепторный потенциал палочек и колбочек
Специфической особенностью фоторецепторов является темповой ток катионов через открытые мембранные каналы внешних сегментов (рис. 17.8). Эти каналы открываются при высокой концентрации циклического гуано- зинмонофосфата, который является вторичным посредником рецепторного белка (зрительного пигмента). Темновой ток катионов деполяризует мембрану фоторецептора до приблизительно —40 мВ, что приводит к выделению медиатора в его синаптическом окончании. Активированные поглощением света молекулы зрительного пигмента стимулируют активность фос- фодиэстеразы — фермента, расщепляющего цГМФ, поэтому при действии света на фоторецепторы в них уменьшается концентрация цГМФ. В результате управляемые этим посредником катионные каналы закрываются, и ток катионов в клетку прекращается. Вследствие непрерывного выхода ионов калия из клеток, мембрана фото  рецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
рецепторов гиперполяризуется приблизительно до —70 мВ, эта гиперполяризация мембраны является рецепторным потенциалом. При возникновении рецепторного потенциала прекращается выделение глутамата в синаптических окончаниях фоторецептора.
Фоторецепторы образуют синапсы с биполярными клетками двух типов, различающихся по способу управления хемозависи- мыми натриевыми каналами в синапсах. Действие глутамата приводит к открытию каналов для ионов натрия и деполяризации мембраны одних биполярных клеток и к закрытию натриевых каналов и гиперполяризации биполярных клеток другого типа. Наличие двух типов биполярных клеток необходимо для формирования антагонизма между центром и периферией рецептивных полей ганглиозных клеток. Адаптация
фоторецепторов к изменениям освещенности
Временное ослепление при быстром переходе от темноты к яркому освещению исчезает спустя несколько секунд благодаря процессу световой адаптации. Одним из механизмов световой адаптации является рефлекторное сужение
зрачков, другой зависит от концентрации ионов кальция в колбочках. При поглощении света в мембранах фоторецепторов закрываются катионные каналы, что прекращает вхождение ионов натрия и кальция и уменьшает их внутриклеточную концентрацию. Высокая концентрация ионов кальция в темноте подавляет активность гуанилатциклазы — фермента, определяющего образование цГМФ из гуанозинтрифосфата. Вследствие снижения концентрации кальция, обусловленного поглощением света, активность гуанилатциклазы повышается, что ведет к дополнительному синтезу цГМФ. Повышение концентрации этого вещества приводит к открытию катионных каналов, восстановлению тока катионов в клетку и, соответственно, способности колбочек отвечать на световые раздражители как обычно. Низкая Концентрация ионов кальция способствует десенситизации колбочек, т. е. уменьшению их чувствительности к свету. Десенситизация обусловлена изменением свойств фосфодиэстеразы и белков катионных каналов, становящихся менее чувствительными к концентрации цГМФ.
Способность различать окружающие предметы исчезает на некоторое время при быстром переходе от яркого света к темноте. Она постепенно восстанавливается в ходе темповой адаптации, обусловленной расширением зрачков и переключением зрительного восприятия с фотопической системы на скотопическую. Темновую адаптацию палочек определяют медленные изменения функциональной активности белков, приводящие к повышению их чувствительности. В механизме темновой адаптации участвуют и горизонтальные клетки, способствующие увеличению центральной части рецептивных полей в условиях низкой освещенности.

Еще по теме Преобразование энергии света в сетчатке:
- ЧУДЕСА СВЕТА И ГРАВИТАЦИИ Парадокс света
- Рецептивные поля клеток сетчатки
- ВЗАИМОДЕЙСТВИЕ СВЕТА С АТОМАМИ И МОЛЕКУЛАМИ
- «Водопад света»
- Неизъяснимые тайны в созерцании Божественного Света Григория Паламы
- 4. Законы движения для квантов света
- ДРЕВНИЙ МИР НОВОГО СВЕТА
- ТРАДИЦИОНАЛИСТСКИЙ МИФ О «КОНЦЕ СВЕТА»
- СУХРАВАРДИ И ФИЛОСОФИЯ СВЕТА
- 11. ПОБОРНИКИ ИСКАЖЕННОГО СВЕТА
- 2. ЭТНОС «ОТРАЖЕННОГО СВЕТА»
Источник

