
Палочки в сетчатке позвоночных
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 18 июля 2018;
проверки требуют 10 правок.
У этого термина существуют и другие значения, см. Палочки.
Сечение слоя сетчатки глаза
Строение палочки сетчатки глаза: 1 — наружный сегмент (содержит мембранные диски), 2 — связующий отдел (ресничка), 3 — внутренний отдел (содержит митохондрии), 4 — основание с нервными окончаниями.
Па́лочки (англ. rod cells) — один из двух типов фоторецепторов, периферических отростков светочувствительных клеток сетчатки глаза, названный так за свою цилиндрическую форму. Это высокоспециализированные клетки, преобразующие световые раздражения в нервное возбуждение. Вторым типом фоторецепторов являются колбочки.
В среднем сетчатка глаза человека содержит около 92 миллионов палочек.[1]
Размеры палочек: длина 0,06 мм, диаметр 0,002 мм.
Палочки чувствительны к свету благодаря наличию в них специфического пигмента — родопсина (или зрительный пурпур). Под действием света происходит ряд очень быстрых превращений и обесцвечивание зрительного пигмента. Чувствительность палочки достаточна, чтобы зарегистрировать попадание даже 2-3 фотонов.
Плотность размещения палочек на различных участках сетчатки глаза неравномерна и может составлять от 20 до 200 тысяч на квадратный миллиметр. Причём на периферии сетчатки их плотность выше, чем к её середине, что определяет их участие в ночном и периферийном зрении. В центре сетчатки, в центральной ямке (жёлтом пятне), палочки практически отсутствуют.
Строение фоторецепторов[править | править код]
Палочки и колбочки сходны по строению и состоят из четырех отделов.
В строении палочки принято различать (см. рисунок):
- Наружный сегмент (содержит мембранные диски с родопсином),
- Связующий отдел (ресничка),
- Внутренний сегмент (содержит митохондрии),
- Область с нервными окончаниями.
В наружном сегменте палочки находится столбик содержащий большое количество мембранных дисков (около тысячи). Мембраны дисков содержат множество молекул светочувствительного пигмента родопсина. Диски представляют собой уплощенные мембранные мешочки, уложенные в виде стопки. Обращённая к свету, наружная часть столбика из дисков, постоянно обновляется, за счет фагоцитоза «засвеченных» дисков клетками пигментного эпителия, и постоянного образования новых дисков, в теле фоторецептора. Диски в колбочке постоянно обновляются (до сотни дисков в сутки). На полное обновление всех дисков фоторецептора требуется около 10 дней.
Внутренний сегмент — это область активного метаболизма, она заполнена митохондриями, поставляющими энергию для обеспечения процессов световосприятия, и полирибосомами, на которых синтезируются белки, участвующие в образовании мембранных дисков и зрительного пигмента. В этом же участке палочки располагается ядро.
К одному интернейрону, собирающему сигнал c сетчатки, как правило, подсоединяются несколько палочек, что дополнительно увеличивает чувствительность глаза (конвергенция). Такое объединение палочек в группы делает периферийное зрение очень чувствительным к движениям и отвечает за феноменальные способности отдельных индивидов к зрительному восприятию событий лежащих вне угла их зрения.
Палочки обладают интересной особенностью. В связи с тем, что все палочки содержат один и тот же светочувствительный пигмент — родопсин, их спектральная характеристика сильно зависит от уровня освещения. При слабом освещении, максимум поглощения родопсина составляет около 500 нм. (спектр сумеречного неба), при этом палочки ответственны за ночное зрение, когда цвета предметов неразличимы. При высоком уровне освещения, родопсин выцветает, его чувствительность падает, и максимум поглощения смещается в синюю область, что позволяет глазу, при достаточном освещении, использовать палочки как приёмник коротковолновой (синей) части спектра[2]. Доказательством того, что приёмником синей части спектра в глазу является палочка, может служить и тот факт, что при цветоаномалии третьего типа (тританопия), глаз человека не только не воспринимает синюю часть спектра, но и не различает предметы в сумерках (куриная слепота), а это указывает именно на отсутствие нормальной работы палочек. Сторонники трёхкомпонентных теорий объяснить эту закономерность до сих пор не могут (почему всегда, одновременно с прекращением работы синего приёмника, перестают работать и палочки).
Таким образом, при ярком свете, палочки совместно с колбочками (которые чувствительны к жёлто-зелёной и жёлто-красной частям спектра)[3] позволяют глазу различать и цвета окружающего нас мира.
Цветное зрение[править | править код]
Нормализованные графики чувствительности человеческих клеток-колбочек различных видов (К, С, Д) и клеток-палочек (П) к различным частям спектра. NB: ось длин волны на данном графике логарифмическая.
Палочки чувствительны в изумрудно-зеленой части спектра (максимум — 498 нм). В остальных частях спектра чувствительны колбочки разных видов. Наличие палочек и разных видов колбочек даёт человеку цветное зрение.
Длинноволновые и средневолновые колбочки (с пиками в жёлто-красном и сине-зелёном диапазонах) имеют широкие зоны чувствительности со значительным перекрыванием, поэтому колбочки определённого типа реагируют не только на свой цвет; они лишь реагируют на него интенсивнее других.[4]
В ночное время, когда поток электромагнитных волн недостаточен для нормальной работы колбочек, зрение обеспечивают только палочки, поэтому ночью человек не может различать цвета.
См. также[править | править код]
- Анкирин 3
- Колбочки
Примечания[править | править код]
- ↑ Curcio, C. A.; Sloan, K. R. et al. Human photoreceptor topography (англ.) // The Journal of Comparative Neurology (англ.)русск. : journal. — 1990. — Vol. 292, no. 4. — P. 497—523. — doi:10.1002/cne.902920402. — PMID 2324310.
- ↑ С. Д. Ременко, «Цвет и зрение», «Картеа Молдовеняскэ», Кишинёв, 1982 г.
- ↑ W. B. Marks, W. U. Dobelle, E. F. Mac Nichol. «Science», v 143, 1964, p 1181.
- ↑
Д. Хьюбел. Глаз, мозг, зрение. — под ред. А. Л. Бызова. — М.: Мир, 1990. — 172 с.
Источник
→ Сетчатка

![]()
рис. 1. Строение палочки (сетчатка)
Палочки (англ. rod cells) — один из двух типов фоторецепторов (экстерорецепторов), периферических отростков светочувствительных клеток сетчатки глаза, названный так за свою цилиндрическую форму. Это высокоспециализированные клетки, преобразующие световые раздражения в нервное возбуждение (сигналы).
Палочки чувствительны к свету благодаря наличию в них специфического пигмента — родопсина (или зрительный пурпур). Под действием света происходит ряд очень быстрых превращений и обесцвечивание зрительного пигмента. В сетчатке глаза у взрослого человека содержится приблизительно около ~90 миллионов палочек[1]. Размеры их очень невелики: длина палочек 0,06 мм, диаметр 0,002 мм. Плотность размещения палочек на различных участках сетчатки глаза неравномерно и может составлять от 20 до 200 тысяч на квадратный миллиметр. В центре сетчатки, в центральной ямке фовеальной области (жёлтом пятне), палочки практически отсутствуют, затем их плотность возрастает и достигает максимальной плотности порядка 140 000-160 000/мм2 в кольце, на расстоянии около 5 мм от центральной ямки. Затем плотность палочек падает до постоянного уровня около 70 000-80 000/мм2 перед тем, как резко уменьшиться на дальней периферии сетчатки. Такое распределение палочек в сетчатке определяет их участие в ночном и периферийном зрении. Чувствительность палочки достаточна, чтобы зарегистрировать попадание даже единичных фотонов.
В ночное время, когда поток фотонов недостаточен для нормальной работы колбочек, зрение обеспечивают только палочки, поэтому ночью человек не может различать цвета.
[править] Строение и исследование фоторецепторов
[править] Морфология
Колбочки и палочки сходны по строению и состоят из четырех участков.
- НАРУЖНЫЙ СЕГМЕНТ (содержит мембранные диски с родопсином),
- СВЯЗУЮЩИЙ ОТДЕЛ (соединительные реснички),
- ВНУТРЕННИЙ СЕГМЕНТ (содержит митохондрии),
- ОБЛАСТЬ С НЕРВНЫМИ ОКОНЧАНИЯМИ (Синаптическая область).
В наружном сегменте палочки находится большое количество мембранных дисков (около тысячи). Мембрана дисков содержит множество молекул пигмента родопсина, которые представляют собой уплощенные мембранные мешочки уложенные в виде стопки монет. Каждый диск образован двумя соединёнными по краям мембранами толщиной порядка 50 — 75 Ангстрем, разделённых промежутком — около 50 Ангстрем.[2].[3]. Диски в колбочке постоянно обновляются (примерно до сотни дисков в сутки).
Связующий отдел содержит 9 пар двойных нитей (фибрилл). Они отходят в соединительной ресничке от одной из двух центриолей (базальное тельце), которые лежат рядом перпендикулярно друг другу. Нити, соединяющие реснички, проходят от внутреннего сегмента до середины наружнего сегмента.[4].
Во внутреннем сегменте имеется скопление радиально ориентированных и плотно упакованных митохондрий. При освещении палочки митохондрии набухают и, вероятно, при этом в них повышается активность окислительных ферментов. Это область активного метаболизма. Митохондрии и полирибосомы, поставляют энергию для обеспечения процессов световосприятия, при этом синтезируются белки, участвующие в образовании мембранных дисков и зрительного пигмента. В этом же участке располагается ядро.[5].[6].
К области с нервными окончаниями палочки подходят и вдаются в неё дендриды биполярных и горизонтальных клеток сетчатки. Кроме того, описаны контакты между рецепторами (палочками и колбочками) сетчатки. В пресинаптических окончаниях обнаружено большое количество синаптических пузырьков (везикул), которые содержат медиатор. Число и размер этих пузырьков, по-видимому, меняются при изменении освещения.[7].[8].[9].
К одному интернейрону, собирающему сигнал c сетчатки, как правило, подсоединяются несколько палочек, что дополнительно увеличивает чувствительность глаза. Такое объединение палочек в группы делает периферийное зрение очень чувствительным к движениям и отвечает за феноменальные способности отдельных индивидов к зрительному восприятию событий лежащих вне угла их зрения.
[править] Процессы происходящие в палочке
В наружных сегментах палочек родопсин ориентирован так, что карбокси-концевой хвост расположен в междисковом (цитоплазматическом) пространстве. Он удерживает 11-цис ретиналь шиффовским взаимодействием с лизиновым остатком в центре «бочки из клепок». При взаимодействии фотона соответствующей длины волны (400 — 600 нм) с 11-цис ретиналем, появляется энергия активации, необходимая для внутримолекулярной перестройки, ведущей к низкоэнергетической транс-форме. Это ключевой момент фоторецепции, все последующее зависит от этого фотохимического преобразования. Транс- ретиналь не может оставаться соединенным с опсином и диффундирует из «бочки», оказываясь в конце концов в пигментном эпителии. Утратив 11-цис ретиналь, молекула опсина претерпевает некоторые конформационные изменения. В этом активированном состоянии она способна реагировать с системой G-белков мембраны наружного сегмента. Эти G-белки известны как трансдуциды или Т-белки. Реакция активированного опсина с альфа-субъединицей Т-белка происходит, как полагают, через его внутриклеточную петлю (в междисковом пространстве), которое приводит к обычному биохимическому процессу. Существует однако и различие — вместо взаимодействия G-белка с аденилатциклазой, которая генерирует цАМФ (обычный случай), альфа-ГТФ- субъединица воздействует на большой тетрамерный фермент цГМФ-фосфодиэстеразу (цГМФ-ФДЭ). Этот мембранно-связанный фермент состоит из альфа-, бета- и двух гамма- субъединиц. Когда этот фермент входит в контакт со свободным Т-альфа-белком, две гамма- субъединицы отделяются. Это растормаживает каталитическую активность альфа- и бета-субъединиц, которые преобразуют цГМФ в 5′-ГМФ. Одновременно две гамма-субъединицы катализируют дефосфорилирование Т-ГТФ в Т-ГДФ. После этого Т-альфа-субъединица отсоединяется от альфа- и бета- субъединиц цГМФ-ФДЭ, что позволяет гамма-субъединицам снова присоединиться с последними. Это приводит к прекращению каталитической активности этого фермента.
[править] Спектральная характеристика палочек
Палочки сетчатки содержат один светочувствительный пигмент — родопсин, их спектральная характеристика сильно зависит от уровня освещения. При слабом освещении, максимум поглощения родопсина составляет около 500 нм. (спектр сумеречного неба), при этом палочки ответственны за сумеречное зрение, когда цвета предметов неразличимы. При высоком уровне освещения, родопсин выцветает, его чувствительность падает, и максимум поглощения смещается в синюю область спектра.
Это свойство палочек подтверждает Эффект Пуркинье, суть которого заключается в том, что при наступлении сумерек, когда освещённость падает, красные цвета чернеют, а белые кажутся голубоватыми. Это объясняется тем, что палочки видят синий край спектра лучше, чем колбочки, но зато колбочки видят, например, тёмно красный цвет, тогда как палочки его совершенно не могут увидеть. [10]
[править] Зрительный фотопигмент палочек — родопсин
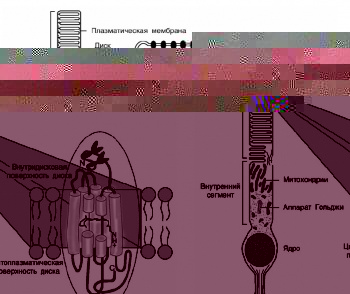
![]()
Рис.2. Схематическое изображение палочки, фоторецепторного диска наружного сегмента, фоторецепторной мембраны диска и молекулы родопсина, в центре которой находится ее хромофорная группа — 11-цис-ретиналь, ковалентно связанный с белковой частью (опсином).[11]
Зрительные фотопигменты опсины в сетчатке позвоночных в фоторецепторах колбочках и палочках (например, родопсин в палочках) — это сложные молекулы, которые представляют собой хромогликопротеиды, содержащие одну хромофорную группу, две олиго-сахаридные цепочки и водонерастворимый мембранный белок опсин. Родопсин является первым мембранным белком животного происхождения, полная аминокислотная последовательность которого была расшифрована в начале 80-х годов Овчинниковым и сотрудниками, Харгрэйвом и сотрудниками (Овчинников и др., 1982, Hargrave et al., 1982). Сравнительно недавно удалось кристаллизовать родопсин, а методом рентгено-структурного анализа в начале с разрешением в 2,8Å, а позже с разрешением в 2,2Å получить его трехмерную структуру (Palczewskiet al., 2000; Liang et al., 2003; Okada et al., 2004)
В сетчатке позвоночных родопсин содержится в фоторецепторах ночного и сумеречного зрения — палочках.
Благодаря приведенным данным в литературе подробно описан хромофорный центр родопсина: конформационное состояние 11-цисретиналя и его взаимодействие с окружающими аминокислотными остатками.
Хромофорной группой всех без исключения зрительных фотопигментов человека и животных является альдегид витамина А1 или витамина А2 или, соответственно, ретиналь1 или ретиналь2, причем только одна из его изомерных форм, а именно его 11-цис-форма. Родопсин — сравнительно небольшой белок: молекулярная масса которого составляет около 40 кДа, а полипептидная цепь состоит из 348 аминокислотных остатков. В молекуле родопсина можно выделить внутримембранный, внутридисковый и цитоплазматический домены. Внутримембранный домен состоит из хромофорного центра и семи трансмембранных α-спиралей, представляющих собой остов опсина. Хромофорная группа — 11-цис-ретиналь ковалентно связан с ε-аминогруппой лизина (Lys-296) спирали ТМ7 через протонированное Шиффово основание. Протонирование Шиффова основания увеличивает делокализацию электрона вдоль полиеновой цепи ретиналя.
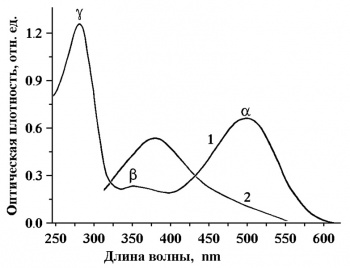
Спектр поглощения родопсина состоит из трех основных полос:
- α- 498 нм,
- β- 350 нм,
- γ- 280 нм (рис.3).
Две первые связаны с поглощением хромофорной группы, а γ-полоса обусловлена в основном поглощением ароматических аминокислот белка — триптофана, тирозина и фенилаланина.
Именно α- полоса в спектре поглощения родопсина определяет кривую видности палочкового сумеречного (скотопического) зрения с максимумом в сине-зеленой области спектра (≈500 нм). Фоточувствительность родопсина необычайно высока. Квантовый выход фотореакции составляет 0,67.
До сих пор остаётся неясным, почему скорость фотоизомеризации ретиналя как хромофора увеличивается почти на два порядка по сравнению со скоростью его фотоизомеризации в растворе. В любом случае роль белкового окружения в этих процессах находится вне сомнения, но внутримолекулярные механизмы, объясняющие этот феномен, остаются до конца не изученными.
[править] Заболевания, связанные с дефектами или отсутствием палочек
- Пигментный ретинит
- Никталопия (общее наименование «ночной слепоты»)
[править] См. также
- Колбочки (сетчатка)
- Ретиномоторная реакция фоторецепторов сетчатки глаза
- Экстерорецепторы
- Палочки в нелинейной теории зрения
[править] Источники
- ↑ Curcio, CA.; Sloan, KR.; Kalina, RE.; Hendrickson, AE. (Feb 1990). «Human photoreceptor topography.». J Comp Neurol 292 (4): 497—523. doi:10.1002/cne.902920402. PMID 2324310.
- ↑ Wolken J. J. 1966. Vision, Thomas C. C.
- ↑ De Robertis E. I., A. Lasansky. 1961. In^ The strukture of the Eye. New York – London : 29
- ↑ Wald G., P. K. Brown, J. R/ Gibbons. 1963. Jur. Opt. Soc. Amer., 53 : 20
- ↑ Островский М. А. 1961. Жур. Общей биол., 22 : 471
- ↑ Лукашевич Т. П. 1962. Доклад АН СССР, 145 : 669
- ↑ Островский М. А. 1961. Жур. Общей биол., 22 : 474
- ↑ Вальцев В. Б. 1965. Жур. Высш. Нервн. Деят., 5 : 934
- ↑ Вальцев В. Б. 1966. Жур. Высш. Нервн. Деят., 16 : 535
- ↑ Ричард Филлипс Фейнман. Фейнмановские Лекции по Физике, том № 3, гл. 35 (Цветовое зрение), стр. 157.
- ↑ https://www.library.biophys.msu.ru/PDF/3353.pdf М. А. Островский ФОТОБИОЛОГИЧЕСКИЙ ПАРАДОКС ЗРЕНИЯ
Источник
Фоторецепторы — светочувствительные сенсорные нейроны сетчатки глаза. Фоторецепторы содержатся во внешнем зернистом слое сетчатки. Фоторецепторы отвечают (а не , как другие нейроны) в ответ на адекватный этим рецепторам сигнал — свет. Фоторецепторы размещаются в сетчатке очень плотно, в виде шестиугольников (гексагональная упаковка)[1][2][3][4].
Классификация фоторецепторов[править | править код]
Maurolicus muelleri
К фоторецепторам в сетчатке глаза человека относятся 3 вида колбочек (каждый тип возбуждается светом определённой длины волны), которые отвечают за цветное зрение, и один вид палочек, который отвечает за сумеречное зрение. В сетчатке глаза человека насчитывается 110 ÷ 125 млн палочек и 4 ÷ 7 млн колбочек[5].
У глубоководной морской рыбы Maurolicus muelleri[en] фоторецепторы дополнены «палочковидными колбочками» («палочкоколбочками», англ. rod-like cones), объединяющими свойства палочек и колбочек и предназначенные для острого зрения при умеренном освещении[6][7].
Сравнение палочек и колбочек[править | править код]
Таблица, иллюстрирующая различия между палочками и колбочками (по книге Эрика Канделя «Принципы науки о нейронах»[8])
| Палочки | Колбочки |
|---|---|
| Используются для ночного зрения (в условиях слабой освещенности) | Используются для дневного зрения (в условиях высокой освещенности) |
| Высокочувствительны; воспринимают и рассеянный свет | Не очень чувствительны к свету; реагируют только на прямой свет |
| Повреждение вызывает никталопию (гемералопию) | Повреждение вызывает слепоту, дневную слепоту, ахроматопсию |
| Низкая острота зрения | Высокая острота зрения; лучшее пространственное разрешение |
| Нет в центральной ямке | Сосредоточены в центральной ямке |
| Замедленная реакция на свет | Быстрая реакция на свет, могут воспринимать более быстрые изменения у раздражителя |
| Имеют больше пигмента, чем колбочки | Имеют меньше пигмента |
| Мембранные диски не привязаны непосредственно к клеточной мембране | Мембранные диски крепятся к наружной мембране |
| В 20 раз больше, чем колбочек, по количеству. | |
| Один тип фоточувствительного пигмента | Три типа фоточувствительных пигментов у человека |
| Ср. Ахроматическое зрение | Ср. Цветное зрение |
Связи между фоторецепторами[править | править код]
У позвоночных животных существуют горизонтальные связи между однотипными фоторецепторами (например, между колбочками с одинаковой чувствительностью), а в некоторых случаях — и между рецепторами разного типа[9][10][11]. В сетчатке приматов связей между палочками не обнаружено[12]. Несмотря на это, фоторецепторы на их освещение отвечают так, будто между ними есть связи. При освещении одного рецептора происходит его гиперполяризация. Если бы не было связей между фоторецепторами, то такое воздействие давало бы единственный отреагировавший фоторецептор сетчатки человека. Однако, опыты показывают, что соседние рецепторы тоже гиперполяризируются. Вероятное объяснение этого парадокса состоит в том, что колбочки центральной ямки расположены очень плотно, и изменение мембранного потенциала одного фоторецептора перетекает на соседние.
См. также[править | править код]
- Глазки Гессе
Примечания[править | править код]
- ↑ Хьюбел Д. Глаз, мозг, зрение. — М.: Мир, 1990. — 240 с.
- ↑ Меденников П. А., Павлов Н. Н. Гексагональная пирамида как модель структурной организации зрительной системы // Сенсорные системы. — 1992. — т.6 № 2 — с.78-83.
- ↑ Лебедев Д. С., Бызов А. Л. Электрические связи между фоторецепторами способствуют выделению протяженных границ между разнояркими полями // Сенсорные системы. — 1988. — т.12, № 3. — с. 329—342.
- ↑ Watson A. B., Ahumada A. J. A hexahonal orthogonal-oriented pyramid as a model of image representation in visual cortex// IEEE Transactions on Biomedical Engineering. — Vol. 36, № 1 — pp.97-106.
- ↑ Измайлов И. А., Соколов Е. Н., Чернорызов А. М. Психофизиология цветового зрения. — М.: Изд-во Московского университета, 1989. — 206 с.
- ↑ de Busserolles F. et al. Pushing the limits of photoreception in twilight conditions: The rod-like cone retina of the deep-sea pearlsides : [англ.] // Science Advances. — 2017. — Vol. 3, no. 11. — P. 1—12 (eaao4709). — doi:10.1126/sciadv.aao4709.
- ↑ У глубоководной рыбки нашли новый тип зрительных рецепторов — «палочкоколбочки», Индикатор. Дата обращения 14 декабря 2017.
- ↑ Kandel, E. R.; Schwartz, J.H.; Jessell, T.M. Principles of Neural Science (неопр.). — 4th. — New York: McGraw-Hill Education, 2000. — С. 507—513. — ISBN 0-8385-7701-6.
- ↑ Школьник-Яррос Е. Г. , Калинина А. В. Нейроны сетчатки. — М.: Наука, 1986. — 208 с.
- ↑ Измайлов И. А., Соколов Е. Н., Чернорызов А. М. Психофизиология цветового зрения. — М.: Изд-во Московского университета, 1989. — 206 с
- ↑ Ноздрачев А. Д. Общий курс физиологии человека и животных. Т.1, — М.: Высшая школа, 1991. −512 с.
- ↑ Hoyenga K. B., Hoyenga K. T. Psychobiology: the neuron and behavior. — Western Illinois University.: Brooks/ Cole Publishing Company Pacific Grove, California, 1988.
Ссылки[править | править код]
- Особенности цветного зрения у различных млекопитающих
Источник

