
Нейронные связи в сетчатке
Элементы нейронной сети сетчатки и их функции
Нейронная сеть сетчатки включает 4 типа нервных клеток:
· ганглиозные клетки,
· биполярные клетки,
· амакриновые клетки,
· горизонтальные клетки.
Ганглиозные клетки – нейроны, аксоны которых в составе зрительного нерва выходят из глаза и следуют в ЦНС. Функция ганглиозных клеток – проведение возбуждения из сетчатки в ЦНС.
Биполярные клетки соединяют рецепторные и ганглиозные клетки. От тела биполярной клетки отходят два разветвленных отростка: один отросток образует синаптические контакты с несколькими фоторецепторными клетками, другой – с несколькими ганглиозными клетками. Функция биполярных клеток – проведение возбуждения от фоторецепторов к ганглиозным клеткам.
Горизонтальные клетки соединяют расположенные рядом фоторецепторы. От тела горизонтальной клетки отходит несколько отростков, которые образуют синаптические контакты с фоторецепторами. Основная функция горизонтальных клеток – осуществление латеральных взаимодействий фоторецепторов.
Амакриновые клетки расположены подобно горизонтальным, но их образуют контакты не с фоторецепторными, а с ганглиозными клетками.
Рис. 2.5. Нейронная сеть сетчатки (А) и электрические ответы разных типов клеток на световой стимул (Б).
Распространение возбуждения в сетчатке
При освещении фоторецептора в нем развивается рецепторный потенциал, который представляет собой гиперполяризацию. Рецепторный потенциал, возникший в фоторецепторной клетке, передается биполярным и горизонтальным клеткам через синаптические контакты с помощью медиатора.
В биполярной клетке может развиваться как деполяризация, так и гиперполяризация (подробнее см. ниже), которая через синаптический контакт распространяется на ганглиозные клетки. Последние являются спонтанно активными, т.е. непрерывно генерируют потенциалы действия с определенной частотой. Гиперполяризация ганглиозных клеток приводит к снижению частоты нервных импульсов, деполяризация – к ее увеличению.
Электрические реакции нейронов сетчатки
Рецептивное поле биполярной клетки представляет собой совокупность фоторецепторных клеток, с которым она образует синаптические контакты. Под рецептивным полем ганглиозной клетки понимают совокупность фоторецепторных клеток, с которыми данная ганглиозная клетка соединена через биполярные клетки.
Рецептивные поля биполярных и ганглиозных клеток имеют круглую форму. В рецептивном поле можно выделить центральную и периферическую часть. Граница между центральной и периферической часть рецептивного поля является динамичной и может смещаться при изменении уровня освещенности.
Реакции нервных клеток сетчатки при освещении фоторецепторов центральной и периферической части их рецептивного поля, как правило, противоположны. При этом существует несколько классов ганглиозных и биполярных клеток (ON-, OFF-клетки), демонстрирующих разные электрические ответы на действие света.
Рис. 2.6. Электрические реакции ганглиозной клетки ON-типа на освещение центральной и периферической части ее рецептивного поля узким пучком света.
1 — ганглиозная клетка, 2 и 3 — центральная и периферическая часть ее рецептивного поля (показано схематично), 4 – пучок света, 5 – отметка времени действия светового стимула, 6 — импульсная электрическая активность ганглиозной клетки
Таблица 2. Классы ганглиозных и биполярных клеток и их электрические реакции
| Классы клеток | Реакция нервных клеток при освещении фоторецепторов, находящихся | |
| в центральной части РП | в периферической части РП | |
| Биполярные клетки ON типа | деполяризация | гиперполяризация |
| Биполярные клетки OFF типа | гиперполяризация | деполяризация |
| Ганглиозные клетки ON типа | Деполяризация и увеличение частоты ПД | Гиперполяризация и снижение частоты ПД |
| Ганглиозные клетки OFF типа | Гиперполяризация и снижение частоты ПД | Деполяризация и увеличение частоты ПД |
| Ганглиозные клетки ON-OFF типа | Дают короткий ON-ответ на стационарный световой стимул и короткую OFF-реакцию на ослабление света. |
Обработка зрительной информации в ЦНС
Сенсорные пути зрительной системы
Миелиновые аксоны ганглиозных клеток сетчатки направляются в головной мозг в составе двух зрительных нервов. Правый и левый зрительные нервы сливаются у основания черепа, образуя зрительный перекрест (хиазму). Здесь нервные волокна, идущие от медиальной половины сетчатки каждого глаза переходят на контрлатеральную сторону, а волокна от латеральных половин сетчаток продолжаются ипсилатерально.
После перекреста аксоны ганглиозных клеток в составе зрительного тракта следуют в латеральные коленчатые тела (ЛКТ), где образуют синаптические контакты с нейронами ЦНС. Аксоны нервных клеток ЛКТ в составе т.н. зрительной лучистости достигают нейронов первичной зрительной коры (поле 17 по Бродману). Далее по внутрикорковым связям возбуждение распространяется во вторичную зрительную кору (поля 18б 19) и ассоциативные зоны коры.
Сенсорные пути зрительной системы организованы по ретинотопическому принципу – возбуждение от соседних ганглиозных клеток достигает соседних точек ЛКТ и коры. Поверхность сетчатки как бы проецируется на поверхность ЛКТ и коры.
Рис. 2.7. Проводящие пути зрительной сенсорной системы.
Х — хиазма, ЛКТ – латеральное коленчатое тело, ЗК – зрительная кора.
Большая часть аксонов ганглиозных клеток заканчиваются в ЛКТ, часть же волокон следует в верхние бугры двухолмия, гипоталамус, претектальную область ствола мозга, ядро зрительного тракта.
· Связь между сетчаткой и верхними буграми четверохолмия служит для регуляции движений глаз.
· Проекция сетчатки в гипоталамус служит для сопряжения эндогенных циркадных ритмов с суточными колебаниями уровня освещенности.
· Связь между сетчаткой и претектальной областью ствола исключительно важна для регуляции просвета зрачка и аккомодации.
· Нейроны ядер зрительного тракта, которые также получают синаптические входы от ганглиозных клеток, связаны с вестибулярными ядрами ствола мозга. Эта проекция позволяет оценивать положение тела в пространстве на основании зрительных сигналов, а также служит для осуществления сложных глазодвигательных реакций (нистагм).
Обработка зрительной информации в ЛКТ
1. Нейроны ЛКТ имеют рецептивные поля округлой формы. Электрические реакции этих клеток аналогичны таковым ганглиозных клеток.
2. В ЛКТ существуют нейроны, которые возбуждаются при наличии в их рецептивном поле границы свет/темнота (контрастные нейроны) или при передвижении этой границы в пределах рецептивного поля (детекторы движения).
Обработка зрительной информации
в первичной зрительной коре
В зависимости от реакции на световые стимулы нейроны коры подразделяют на несколько классов.
Нейроны с простым рецептивным полем. Наиболее сильное возбуждение такого нейрона происходит при освещении его рецептивного поля световой полоской определенной ориентации. Частота нервных импульсов, генерируемых таким нейроном уменьшается при изменении ориентации световой полоски (рис. 2.8 А).
Рис. 2.8. Имульсация различных нейрон зрительной коры в ответ на освещение их рецептивных полей различными световыми стимулами.
А – нейрон с простым рецептивным полем; Б – нейрон со сложным рецептивным полем; В – нейрон со сверхсложным рецептивным полем: 1 – периферическая часть рецептивного поля, 2 – центральная часть рецептивного поля, 3 – световой стимул.
Нейроны со сложным рецептивным полем. Максимальная степень возбуждения нейрона достигается при передвижении светового стимула в пределах ON зоны рецептивного поля в определенном направлении. Передвижение светового стимула в другом направлении или выход светового стимула за пределы ON зоны вызывает более слабое возбуждение (рис. 2.8 Б).
Нейроны со сверхсложным рецептивным полем. Максимальное возбуждение такого нейрона достигается при действии светового стимула сложной конфигурации. Например, известны нейроны, наиболее сильное возбуждение которых развивается при пересечении двух границ между светлым и темным в пределах ON зоны рецептивного поля (рис. 2.8 В).
Несмотря на огромно количество экспериментальных данных о закономерностях реагирования клеток на разные зрительные стимулы, к настоящему времени нет полной теории, объясняющей механизмы обработки зрительной информации в головном мозге. Мы не можем объяснить, каким образом разнообразные электрические реакции нейронов сетчатки, ЛКТ и коры обеспечивают распознавание образов и другие феномены зрительного восприятия.
Регуляция функций вспомогательного аппарата
Регуляция аккомодации. Изменение кривизны хрусталика осуществляется при помощи цилиарной мышцы. При сокращении цилиарной мышцы кривизна передней поверхности хрусталика увеличивается и преломляющая сила возрастает. Гладкомышечные волокна цилиарной мышцы иннервируются постганглионарными нейронами, тела которых располагаются в цилиарном ганглии.
Адекватным стимулом для изменения степени кривизны хрусталика является нечеткость изображения на сетчатке, которая регистрируется нейронам первичной коры. За счет нисходящих связей коры происходит изменение степени возбуждения нейронов претектальной области, что в свою очередь вызывает активацию или торможение преганглионарных нейронов глазодвигательного ядра (ядро Эдингера–Вестфаля) и постганглионарных нейронов цилиарного ганглия.
Регуляция просвета зрачка. Сужение зрачка происходит при сокращении кольцевых гладкомышечных волокон роговицы, которые иннервируются парасимпатическими постганглионарными нейронами цилиарного ганглия. Возбуждение последних происходит при высокой интенсивности падающего на сетчатку света, которая воспринимается нейронами первичной зрительной коры.
Расширения зрачка осуществляется при сокращении радиальных мышц роговицы, которые иннервируются симпатическими нейронами ВШГ. Активность последних находится под контролем цилиоспинального центра и претектальной области. Стимулом для расширения зрачка является уменьшение уровня освещенности сетчатки.
Регуляция движений глаз. Часть волокон ганглиозных клеток следуют к нейронам верхних бугров четверохолмия (средний мозг), которые связаны с ядрами глазодвигательного, блокового и отводящего нервов, нейроны которых иннервируют поперечнополосатые мышечные волокна мышц глаза. Нервные клетки верхних бугров получат синаптические входы от вестибюлярных рецепторов, проприорецепторов мышц шеи, что позволяет организму координировать движения глаз с перемещениями тела в пространстве.
Феномены зрительного восприятия
Распознавание образов
Зрительная система обладает замечательной способность распознавать объект при самых разных вариантах его изображения. Мы можем узнавать образ (знакомое лицо, букву и т. п.), когда некоторых его частей недостает, когда он содержит лишние элементы, когда он по-разному ориентирован в пространстве, имеет разные угловые размеры, повернут к нам разными сторонами и т.п. Нейрофизиологичекие механизмы этого феномена в настоящее время интенсивно изучаются.
Рис. 2.9. Распознавание образов: человек легко распознает знакомые буквы при их разной ориентации, разном варианте написания, разных размерах, отсутствии части изображения и даже может составить образ из разрозненных фрагментов.
Постоянство формы и размеров
Как правило, мы воспринимаем окружающие предметы неизменными по форме и размерам. Хотя на самом деле их форма и размеры на сетчатке не являются постоянными. Например, велосипедист в поле зрения всегда кажется одинаковым по величине независимо от расстояния до него. Колеса велосипеда воспринимаются как круглые, хотя на самом деле их изображения на сетчатке могут быть узкими эллипсами. Это явление демонстрирует роль опыта в видении окружающего мира. Нейрофизиологические механизмы этого феномена в настоящее время неизвестны.
Восприятие глубины пространства
Изображение окружающего мира на сетчатке является плоским. Однако, мы видим мир объемным. Существует несколько механизмов, которые обеспечивают построение 3-мерного пространства на основании плоских изображений, сформированных на сетчатке.
· Поскольку глаза расположены на некотором расстоянии друг от друга, то изображения, формирующиеся на сетчатке левого и правого глаза, несколько различаются друг от друга. Чем ближе расположен объект по отношению к наблюдателю, тем больше будут различаться эти изображения.
· Чем дальше расположен объект, тем меньше угловой размер его изображения на сетчатке. Оценивая угловые размеры знакомых предметов, мы можем оценивать расстояние до них.
· Перекрывание изображений также помогает оценить их взаимное расположение в пространстве. Изображение близкого предмета может перекрывать изображение удаленного, но не наоборот.
· При смещении головы наблюдателя изображения наблюдаемых объектов на сетчатке также будут смещаться (явление параллакса). При одном и том же смещении головы изображения близких объектов будут смещаться сильнее, чем изображения удаленных
Источник
Сетчатка состоит из наружного пигментного и внутреннего светочувствительного слоёв. Внутренний её слой имеет зрительную (заднюю) и слепую (переднюю) части. На задней поверхности сетчатки находится слепое пятно — место выхода зрительного нерва. Несколько латеральнее от него располагается желтое пятно — участок наилучшего зрения. Сетчатка представляет цепь нейронов, между которыми находятся глиоциты и сосуды (рис. 23).
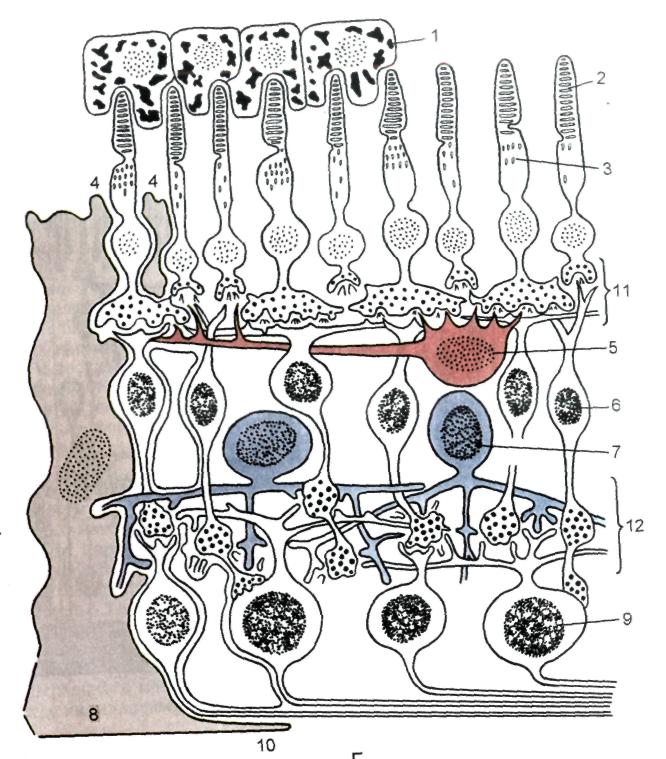
Рис. 23. Схема строения сетчатки. 1 – пигментоциты; 2 – палочки; 3 – колбочки; 4 – зона расположения наружной пограничной (глиальной) мембраны; 5 – гоРИзонтальные клетки; 6 – биполярные (вертикальные) клетки; 7 – амакринные клетки; 8 – глиоцит (мюллеровы волокна); 9 – ганглиозные клетки; 10 – зона расположения внутренней пограничной (глиальной) мембраны; 11 и 12 – синапсы.
В состав нейронов сетчатки входят светочувствительные (фоторецепторные), биполярные (ассоциативные) и ганглиозные нейроны, которые, располагаясь радиально, связаны между собой синапсами. Кроме этого, есть ещё два вида нейронов, обеспечивающих связь по горизонтали: горизонтальные клетки (соединяют фоторецепторные и биполярные нейроны) и амакринные клетки (соединяют биполярные и ганглиозные нейроны).
Фоторецепторные нейроны — типичные биполярные клетки. Их периферические отростки – дендриты имеют вид палочек и колбочек и образуют одноимённый слой. Центральные отростки – аксоны — образуют синаптические связи с биполярными и горизонтальными клетками.
И палочковые, и колбочковые отростки состоят из наружного и внутреннего сегментов, связанных ресничкой. Наружные сегменты располагаются между отростками клеток пигментного эпителия.
ПалочковИднЫе нейроны располагаются в периферических отделах сетчатки, воспринимают световые сигналы низкой интенсивности (сумеречное зрение) и отвечают за черно-белое зрение. Они имеют узкие, вытянутые периферические отростки (палочки), наружный сегмент которых имеет цилиндрическую форму и содержит стопкообразно расположенные мембранные диски (уплощощенные мешочки). В дисках находится зрительный пигмент родопсин. Под влиянием света родопсин расщепляется на составляющие: белок опсин и альдегид витамина А. Это влечёт гиперполяризацию рецепторов и изменение ионной проницаемости мембран, вследствие чего возникает биоэлектрический потенциал. В темноте осуществляется восстановление родопсина.
В проксимальных участках наружных сегментов происходит постоянное обновление дисков и их смещение в дистальные, где по мере старения они фагоцитируются пигментным эпителием. При недостаточном поступлении в организм витамина А обновление дисков нарушается, а при его отсутствии они разрушаются («куриная слепота»).
Внутренний сегмент палочек содержит удлиненные митохондрии, центриоли, элементы АЭС и ГЭС, комплекс Гольджи. Он обеспечивает наружный сегмент энергией и веществами, необходимыми для фоторецепции. Ядро палочковых фоторецепторов мелкое, округлое, окружено тонким ободком цитоплазмы. Аксоны заканчиваются шаровидными утолщениями.
КолбочковИдныЕ нейроны располагаются в центральных отделах сетчатки и особенно многочисленны в центральной ямке желтого пятна, реагируют на свет высокой интенсивности, обеспечивают дневное и цветовое зрение. Наружные сегменты колбочек имеют коническую форму. Они более короткие и широкие, чем у палочковых клеток. Мембранные диски в них образованы складками плазмолеммы и не отделены от неё. В мембране дисков содержится зрительный пигмент йодопсин, который в функционально различных типах колбочек разлагается под действием красного, зеленого или синего света. В колбочках, в отличие от палочек, не происходит постоянного перемещения дисков и их фагоцитоза пигментным эпителием. Строение внутреннего сегмента колбочек сходно с таковым у палочек. Ядра — крупнее и светлее, чем у палочковых клеток. Аксоны заканчиваются в наружном сетчатом слое расширением треугольной формы.
Биполярные (ассоциативные) нейроны своими дендритами образуют синапсы с аксонами фоторецепторных клеток, а аксонами — с дендритами ганглиозных и амакринных клеток. Подразделяются на несколько типов.
ГанглиоЗНые нейроны – самые крупные мультиполярные клетки с эксцентрично расположенным ядром и крупным ядрышком. В цитоплазме содержатся хорошо развитые органеллы. Дендриты образуют синапсы с аксонами биполярных клеток и отростками амакринных клеток, а аксоны образуют зрительный нерв.
Конвергенция (схождение) нервных импульсов в сетчатке обеспечивается характером связей ее нейронов и свойственна всем отделам, за исключением центральной ямки. Несколько палочковых клеток образуют синапсы на одной биполярной, а несколько биполярных контактируют с одной ганлионарной. Общий показатель конвергенции в сетчатке равен 105:1.
Горизонтальные нейроны — ассоциативные мультиполярные клетки; их дендриты и аксон синаптически связаны с аксонами палочковых и колбочковых клеток, а также с дендритами биполярных нейронов.
Амакринные клетки — ассоциативные нейроны, у которых, как правило, аксоны не выявляются, но некоторые из них содержат длинный аксоноподобный отросток. Дендриты образуют связи с аксонами биполярных клеток.
Пигментный эпителий сетчатки располагается на границе с сосудистой оболочкой и своими отростками проникает в фотосенсорный слой. Он образован кубическими (на периферии сетчатки) или призматическими (в ее центре) клетками с базально расположенными ядрами. В цитоплазме хорошо представлены органеллы общего значения и, особенно, лизосомы. Содержат большое количество гранул меланина. Базальная часть плазмолеммы имеет выраженную складчатость — признак активного ионного транспорта. Латеральные поверхности связаны десмосомами. Апикальная поверхность содержит микроворсинки и длинные ветвящиеся отростки, которые располагаются между палочками и колбочками фоторецепторных нейронов.
В функциональном плане пигментный эпителий осуществляет трофику фоторецепторных нейронов, транспорт к их периферическим отросткам витамина А, и др. веществ, способствующих регенерации; фагоцитоз и переваривание кончиков наружных сегментов палочковых фоторецепторов; обеспечение избирательной диффузии веществ из сосудисто-капиллярного слоя сосудистой оболочки; регуляцию поступления светового потока к рецепторам и предотвращение избыточной их засветки, благодаря синтезу и перемещению меланина в отростки на свету. В темноте гранулы меланина перемещаются из отростков в тело клеток.
Нейроглия сетчатки представлена радиальными глиоцитами {мюллеровыми клетками), астроцитами и микроглией.
Мюллеровы клетки пронизывают почти всю толщину сетчатки. Своими основаниями они на границе со стекловидным телом формируют внутреннюю глиальную пограничную мембрану, а апикальными отростками образуют у основания палочек и колбочек наружную глиальную пограничную мембрану. Выполняют поддерживающую и трофическую функции. Вместе с астроцитами они образуют гемато-ретинальный барьер.
Микроглиоциты немногочисленны, располагаются во всех слоях сетчатки, осуществляют фагоцитоз.
Слои сетчатки:
Периферические отростки фотосенсорных нейронов образуют слой палочек и колбочек, ядросодержащие участки клеток — наружный, внутренний ядерные и ганглиозный слои, а области синаптических связей нейронов – наружный и внутренний сетчатые слои. Аксоны ганглиозных нейронов формируют слой нервных волокон, образующих зрительный нерв.
Кроме того, слой палочек и колбочек отделён от наружного ядерного слоя внутренней глиальной пограничной мембраной, а на границе слоя нервных волокон и стекловидного тела располагается наружная глиальная пограничная мембрана.
Кровоснабжение сетчатки осуществляется ветвями центральной артерии сетчатки. Проникая вместе со зрительным нервом, она в области сосочка разделяется на радиально расположенные ветви, которые сначала располагаются между стекловидным телом и внутренней глиальной пограничной мембраной, а затем заходят в слои сетчатки, образуя капиллярное сплетение, доходящее до внутреннего ядерного слоя. Трофика расположенных кнаружи от него слоёв сетчатки осуществляется за счёт сосудисто-капиллярного слоя сосудистой оболочки диффузно через слой пигментного эпителия.
Источник

