
Меланопсинсодержащие ганглиозные клетки сетчатки
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг. Аксоны ганглионарных клеток не миелинизированы при прохождении сетчатки, чтобы не препятствовать прохождению света. Далее они покрыты миелиновой оболочкой.
Ганглионарные клетки завершают «трёхнейронную рецепторно-проводящую систему сетчатки»: фоторецептор — биполярный нейрон — ганглионарная клетка.
Функции ганглионарных клеток[править | править код]
Клетки сетчатки связаны между собой сложной сетью возбуждающих, подавляющих и двунаправленных сигнальных связей. Они собирают информацию от всех слоев сетчатки как по вертикальным путям (фоторецепторы — биполяры — ганглионарные клетки), так и по латеральным путям (фоторецепторы — горизонтальные клетки — биполяры — амакриновые клетки — ганглионарные клетки).
Рецептивные поля[править | править код]
На одну ганглионарную клетку приходится от одного до сотни биполярных нейронов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от нескольких до нескольких тысяч фоторецепторов. Каждая ганглионарная клетка суммирует сигналы от большого числа фоторецепторов, что повышает световую чувствительность, но уменьшает разрешение. Фоторецепторы, соединенные с одной ганглионарной клеткой, образуют её рецептивное поле. Рецептивные поля ганглионарных клеток подразделяются на простые и сложные. Простые поля — имеют концентрическую структуру, подобно полям биполярных нейронов. Сложные — разделяются на несколько различных функциональных зон. Рецептивные поля могут перестраиваться, адаптируясь к уровню освещённости и характеристикам световых стимулов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от единиц до десятков тысяч фоторецепторов (палочек и колбочек). В свою очередь, один фоторецептор через биполярные нейроны может быть связан с десятками ганглионарных клеток. В среднем на 100 фоторецепторных клеток приходится одна ганглионарная (т.е., их от 1.2 до 1.5 млн). Чем ближе к центральной ямке глаза, тем меньше фоторецепторов приходится на одну ганглионарную клетку. Поэтому у людей слабое периферическое зрение. В районе центральной ямки, наоборот, высокое разрешение, но менее высокая светочувствительность, поскольку здесь каждый фоторецептор (колбочка) соединен с одной биполярной (карликовой) клеткой, которая в свою очередь соединена лишь с одной ганглионарной.
Типы ганглионарных клеток[править | править код]
Существует всего 18 типов ганглионарных клеток сетчатки.
Большинство относится к трем типам:
1. Парвоганглионарные клетки — карликовые клетки (около 80% от числа всех ганглионарных клеток сетчатки), имеющие средний размер тела и маленькое дерево дендритов, входят в карликовый путь (чувствительный путь, ведущий от глаза к четверохолмию) и связаны с парвоцеллюлярными (мелкоклеточными) слоями латеральных коленчатых тел. С этими клетками связывают высокую остроту зрения и цветовое зрение.
2. Магноклетки — (около 10%) очень разнообразны (малые и большие зонтичные клетки): с большими телами и многочисленными укороченными ветвями, маленькими телами и большим разветвлением дендритов, которые проецируются в крупноклеточные слои латеральных коленчатых тел. Отвечают за востриятие движущихся объектов. Имеют большие рецепторные поля.
3. Кониоцеллюлярные клетки очень мелкие, составляют от 8 до 10% всех ганглионарных клеток сетчатки. Получают сигналы от среднего количества фоторецепторов. Имеют очень большие рецептивные поля. Всегда ON для колбочек синего цвета и OFF для красного и зеленого.
Выделяют ганглионарные клетки, связанные с палочковыми и колбочковыми нейронами, с on- и off-центрами, которые отвечают на световое раздражение деполяризацией или гиперполяризацией соответственно. Дендриты клеток с on-центром разветвляются в подуровне а (пигментном эпителии?), с off-центром в подуровне G (ганглионарном слое?) внутреннего сетчатого слоя. Цветовой канал связан с красным, зеленым и синим типом on/off-ганглионарных клеток. Если красный и зеленый тип ганглионарных on/off-клеток относится к карликовому пути, то синий тип не относится к последнему. On/off-ответы ганглионарных клеток определяются специальными контактами колбочковых биполяров и расположением ганглионарных клеток в соответствующем подуровне внутреннего сетчатого слоя
Светочувствительные ганглионарные клетки[править | править код]
В 1991 году были открыты особые светочувствительные ганглионарные клетки типа ipRGC (intrinsically photosensitive retinal ganglion cells), или mRGC (melanopsin-containing retinal ganglion cells). Они, в отличие от ранее известных ганглионарных клеток, содержат светочувствительный пигмент меланопсин, отличающийся от других фоточувствительных пигментов глаза: родопсина палочек и йодопсина колбочек. И этим они отличаются от других ганглионарных клеток, находящихся в сетчатке глаза, которые не умеют реагировать непосредственно на свет.
Эти светочувствительные ганглионарные клетки — новый, третий тип фоторецепторов сетчатки глаза, помимо известных уже в течение 200 лет палочек и колбочек. Они напрямую возбуждаются под действием света даже при блокировании «классических» фоторецепторов глаза — палочек и колбочек.
Нервные пути от этих ганглиозных (ганглионарных) клеток ведут порождённое в них светом возбуждение от сетчатки к гипоталамусу тремя разными путями, обеспечивая световое управление циркадными ритмами, а также по отдельному нервному пути обеспечивают реакцию сужения зрачка на свет.
Литература[править | править код]
- Ноздрачёв А. Д., Баженов Ю. И., Баранникова И. А., Батуев А. С. и др. Начала физиологии: Учебник для вузов / Под ред. акад. А. Д. Ноздрачёва. СПб.: Лань, 2001. 1088 с.
Ссылки[править | править код]
- Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System
- Photosensitive ganglion cells
- Vision beyond image formation: The role of melanopsin cells in regulating mammalian physiology
- Blind Mice Can «See» Thanks To Special Retinal Cells
- Фоторецепторы и фоторецепция
[1]
[2]
[3]
[4]
[5]
Источник
Фоторецепторы человека
Фоторецепторами являются палочки и колбочки , расположенные в наружном слое сетчатки . Палочки и колбочки сходны по своему строению, они состоят из четырех участков:
1. Наружный сегмент — светочувствительный участок, где световая энергия преобразуется в рецепторный потенциал . Наружный сегмент заполнен мембранными дисками, образованными плазматической мембраной.
В палочках в каждом наружном сегменте содержится 600 — 1000 дисков, которые представляют собой уплощенные мембранные мешочки, уложенные как столбик монет. В колбочках мембранных дисков меньше, они представляют собой складки плазматической мембраны.
2.
Фоторецепторы: строение и функции. Зрительные пигменты. Строение сетчатки
Перетяжка — место, где наружный сегмент почти полностью отделен от внутреннего впячиванием наружной мембраны. Связь между двумя сегментами осуществляется через цитоплазму и пару ресничек, переходящих из одного сегмента в другой.
3. Внутренний сегмент — область активного метаболизма, заполненная митохондриями, доставляющими энергию для процессов зрения, и полирибосомами, на которых синтезируются белки, участвующие в образовании мембранных дисков и зрительного пигмента.
Здесь же расположено ядро.
4. Синаптическая область — место, где клетка образует синапсы с биполярными клетками. Диффузные биполярные клетки могут образовывать синапсы с несколькими палочками. Это явление, называемое синаптической конвергенцией, уменьшает остроту зрения, но повышает светочувствительность глаза.
Моносинаптические биполярные клетки связывают одну колбочку с одной ганглиозной клеткой, что обеспечивает лучшую по сравнению с палочками остроту зрения. Горизонтальные клетки и амакриновые клетки связывают вместе некоторое число палочек или колбочек.
Благодаря этим клеткам зрительная информация еще до выхода из сетчатки подвергается определенной переработке. Эти клетки участвуют также в латеральном торможении.
Палочек в сетчатке содержится больше, чем колбочек — 120 млн и 6 — 7 млн соответственно.
Тонкие, вытянутые палочки размером 50х3 мкм равномерно распределены по всей сетчатке, кроме центральной ямки, где преобладают удлиненые конические колбочки размером 60х1,5 мкм. Так как в центральной ямке колбочки очень плотно упакованы (150 тыс. на кв.мм), этот участок отличается высокой остротой зрения. Палочки обладают большей чувствительностью к свету и реагируют на более слабое освещение. Палочки содержат только один зрительный пигмент, не могут различать цвета и используются преимущественно в ночном зрении .
Колбочки содержат три зрительных пигмента, что позволяет распознавать цвета, они используются преимущественно при дневном свете. Палочковое зрение отличается меньшей остротой, так как палочки расположены менее плотно и сигналы от них подвергаются конвергенции, но именно это обеспечивает высокую чувствительность, необходимую для ночного зрения.
Палочки содержат светочувствительный пигмент родопсин .
Клетки фоторецепторы:
палочки
колбочки
См. ФОТОРЕЦЕПТОРЫ ЖИВОТНЫХ
See Фоторецептор
Ссылки:
Все ссылки
Фоторецепторы глаза
Описаны три вида фоторецепторов сетчатки глаза: палочки, колбочки и пигментосодержащие ганглиозные клетки.
Рецепторный отдел зрительного анализатора.
Раньше (в течение 200-летней истории исследования глаза) считалось, что рецепторный отдел зрительного анализатора (зрительной сенсорной системы) состоит из фоторецепторов двух типов, но теперь мы должны говорить о трёх типах фоторецепторов сетчатки:
1.Колбочки (их 6-7 млн): им нужна высокая освещенность, они имеют разную чувствительность к разному спектру (длине волны), обеспечивают цветовое зрение, содержат пигмент йодопсин.
2.Палочки (их 110-120 млн): они работают при слабой освещенности, имеют очень высокую чувствительность, но не различают цвета и дают не резкое изображение, содержат пигмент родопсин («зрительный пурпур»).
Эти два типа фоторецепторов расположены в рецепторном слое сетчатки глаза перпендикулярно к направлению светового луча (столбиками).
Причём они, можно сказать, неприлично развёрнуты к свету тылом.
Но относительно недавно в сетчатке были обнаружены фоторецепторы третьего типа:
3. Меланопсинсодержащие ганглиозные клетки сетчатки (МГКС), или же intrinsically photosensitive retinal ganglion cells (ipRGCs): их всего 2% среди ганглиозных клеток сетчатки, они реагируют на освещённость, но не дают зрительных образов, содержат пигмент меланопсин, который сильно отличается от родопсина палочек и йодопсина колбочек.
Нервные пути от этих ганглиозных (ганглионарных) клеток ведут световое возбуждение от сетчатки к гипоталамусу тремя разными путями.
В палочках и колбочках содержатся светочувствительные пигменты. Оба пигмента имеют в своей основе видоизмененный витамин А.
Фоторецепторы: строение и функции. Зрительные пигменты. Строение сетчатки
Если не хватает витамина А, то страдает зрительное восприятие, т.к. не хватает «заготовок» для производства зрительного пигмента.
Палочки имеют максимум поглощения света в области 500 нм.
Колбочки же, в отличие от палочек, бывают трех типов:
1. «Синие» (коротковолновые — S) — 430-470 нм. Их 2% от общего числа колбочек.
2. «Зелёные» (средневолновые — M) – 500-530 нм. Их 32%.
3. «Красные» (длинноволновые — L) – 620-760 нм.
Их 64%.
В каждом виде фоторецепторов используется свой тип зрительного пигмента. Интересно, что в 2000-е годы была обнаружена огромная вариабельность в соотношении красных и зелёных колбочек у разных людей. Стандартное соотношение, приведённое выше, составляет 1:2, но оно может достигать и 1:40, если сравнивать между собой разных людей. И тем не менее мозг компенсирует эти различия, и люди с разным соотношением красных и зелёных колбочек могут одинаково называть цвет с одной длиной волны.

Фотохимические процессы в глазу идут экономно: даже на ярком свету распадается только малая часть пигмента.
В палочках это всего 0,006%. В темноте пигменты восстанавливаются.
Родопсин – пигмент палочек.
Йодопсин – пигмент красных колбочек.
Йодопсин восстанавливается быстрее родопсина в 530 раз, поэтому при недостатке витамина А, в первую очередь страдает зрение палочек, или сумеречное зрение.
Слой фоторецепторов лежит на слое пигментных клеток, которые содержат пигмент фуксин.
Он поглощает свет и обеспечивает чёткость зрительного восприятия.
Отличительная черта фоторецепторов – это не деполяризация, а гиперполяризация в ответ на раздражение.
Можно сказать, что действие света как бы «повреждает» фоторецептор, разрушает его белок, и он перестает нормально работать, впадает в заторможенное состояние.
Фотохимическая «хрупкость» фоторецепторных клеток сетчатки и клеток пигментного эпителия к отоповреждению связана со следующими факторами:
1) присутствием в них эффективно поглощающих свет фотосенсибилизаторов,
2) достаточно высоким парциальным давлением кислорода,
3) наличием легко окисляющихся субстратов, в первую очередь полиненасыщенных жирных кислот в составе фосфолипидов.
Именно поэтому в ходе эволюции органов зрения позвоночных и беспозвоночных сформировалась достаточно надежная система защиты от опасности фотоповреждения (Островский, Федорович, 1987).
Эта система включает постоянное обновление светочувствительных наружных сегментов зрительных клеток, набор антиоксидантов и оптические среды глаза как светофильтры, где ключевую роль играет хрусталик.
Источник

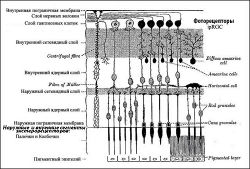
Схема слоёв поперечного сечения сетчатки глаза. Область, помеченная «ганглиозный слой», содержит относящиеся к сетчатке глаза клетки нервного узла клетки фоторецепторы ipRGC
Ошибка создания миниатюры: Файл не найден
Меланопсин — фотопигмент, один из опсинов, непосредственно участвует в зрительном процессе, регуляции циркадных ритмов; он находится в специализированных фоточувствительных ганглиальных клетках сетчатки глаза, в коже и мозговых тканях живонтных. Этот зрительный пигмент (фотопигмент) был обнаружен в ганглиальных клетках ipRGC сетчатки глаза млекопитающих.[2]
Мелаопсин принимает участие в режиме «сна-бодрствования», супрессии эпифизального мелатонина у человека; участвует в зрачковом рефлексе, в формировании зрительных образов.
[3][4]
Рецепторы меланопсина связаны с индивидуальной чувствительностью к свету, а также с болями от мигрени и др.. [5],[6]
Открытие и функции[править]
Меланопсин был обнаружен в 1998 г. в специализированных светочувствительных клетках кожи лягушек (см. работу en:Ignacio Provencio с сотр. [7] В 1999 г. др. Russell Foster показал, что в глазу млекопитающих что существует третий класс фоторецепторов. В 2000 г. др. Provencio показал, что в тканях млекопитающих, включая человека, также синтезируется меланопсин. Он был найден только в небольшом числе ганглиарных клеток, воспринимающих световой сигнал из сетчатки.
Меланопсин принимает разнообразное участие в процессах формирования ритмов «сон-бодрствование» у человека и животных (крысы).
Меланопсин — представитель опсинов[править]
Когда свет активизирует меланопсин, сигнализирующая система, меланопсин-содержащие клетки нейронной сети тормозят прохождение импульсов, которые проводятся через их аксоны к определенным мозговым зонам.
Эти зоны включают olivary pretectal ядро (OPN) (центр, ответственный за управление зрачков глаза) и, через ретиногипоталамический путь (RHT), в suprachiasmatic ядро hypothalamus — центр циркадных ритмов.
Клетки нервного узла, содержа фотопигмент меланопсин, как полагают, влияет на эти цели, выпуская от их терминалов аксона медиаторы — глутаматную и гипофизарную аденилатциклазу, активизирующая многопептид (PACAP), а также получает вход к палочкам и колбочкам, управляет сигналами клеток входа (ДА,НЕТ) к этим тропам.
Мутация гена, связанного с экспрессией меланопсина, приводит к возникновению т.н. «Сезонного Эмоционального нарушения».[8]
Ошибка создания миниатюры: Файл не найден
- Рис. 2. Melanopsin-ген. (a) Определена аминокислотная последовательность и предсказана вторичная структура меланопсина. Заштрихованная область указывает трансмембранные области. (b) Филогенетическое дерево, связывающее меланопсин с др. представителями опсинов позвоночных и нехарактерных разновидностей опсинов (L, чувствительных к длинноволновому участку спектра; M1, синяя-подобная — «средневолновой» опсин; M2, зелено-подобная — «среднволновой» опсин; P, шишковидный опсин; Rh, родопсин; S, «коротковолновой опсин», чувствительны все клетки» (цыпленок); VA, позвоночное животное, древнее опсин атлантического лосося. (c) Hydropathy анализ для вторичного предсказания структуры. (Provencio и др. 1998).
Уникальная способность клеток ipRGCs, чтобы ответить на свет, происходит только за счёт выделенного фотопигмента меланопсина (melanopsin). Первоначально клонированный от лягушки кожный melanophores, melanopsin ген (OPN4) имеется orthologs у многих млекопитающих разновидностях, включая мышей, обезьян, и людей (Provencio, Родригес и др. 2000). Анализ гидрофобной последовательности аминокислоты melanospin предсказывает, что 7-трансмембранная структура, обычная во всех Белках-G , которая объединила рецепторы (рис. 2a, 2c) (Provencio, Jiang и др. 1998). Мкланопсин универсален, он больше похож на беспозвоночный rhabdomeric opsins (r-opsins), чем ресничный opsins разновидностей позвоночных (c-opsins), при чём он больше, в качестве меланопсина может сигнализировать о свете посредством различных механизмов, чем используемые позвоночными палочками и колбочками фотопигменты (рис. 2b) (Provencio, Jiang и др. 1998).
Хотя первые исследования ipRGCs настоятельно предложили меланопсин как фотопигмент в клетках, выполняющий роль для группы клеток синего цвета, поглощающего известным flavoproteins, но только пока cryptochromes не мог быть первоначально исключен (Berson 2007). Функцию Cryptochromes в качестве циркадных фотопигментов у беспозвоночных он быстро одобрил их в сфере функционирования у млекопитающих как циркадные фотопигменты (Kavakli и 2002 Sancar; Ван Джелдер, Gibler и др. 2002). Однако, теперь имеется подавляющее доказательство того, что melanopsin является фотопигментом в клетках ipRGCs, и таким образом является истинным циркадным фотопигментом. Когда melanopsin ген был удален при помощи трансгенных методов в мышах (мыши нокаута), то относящиеся к сетчатке глаза клетки нервного узла, помеченные от SCN, больше не могли сигнализировать свет(рис. 3) (Лукас, Hattar и др. 2003). Кроме того, животные, с введенным melanopsin-геном (мутантом) показывают дефициты в многократных визуальных отражениях, типа сжатия зрачка и восприятия света (Панда, Sato и др. 2002; Рубин, Brennan и др. 2002; Лукас, Hattar и др. 2003; Панда, Provencio и др. 2003).
Как дальнейший тест, Турция и. ал использовал множество плоских мультиэлектродов, которые позволяют делать запись extracellularly от множеств клеток нервного узла сетчатки глаза сразу, позволяя изолировать многократные легкие ответы от клеток ipRGCs. Напрмер, клетки сетчатки глаза мышей нокаута от melanopsin не показали никаких свойственных фотоответов от клеток нервного узла (Турция, Zhang и др. 2005). Несмотря на эти доказательства, все еще были споры относительно способности melanopsin функционировать в качестве истинного фотопигмента. Но споры были твердо погашены на примере ряда изящных экспериментов, где посредством melanopsin-гена было показано, что при многократных опытах на разных клетках, которые являются обычно нечувствительными к свету, они сильно отвечали на свет. Это указывает на способность melanopsin функционировать как bonafide фотопигмент (рис. 3) (Melyan, Tarttelin и др. 2005; Панда, Nayak и др. 2005; Qiu, Kumbalasiri и др. 2005).
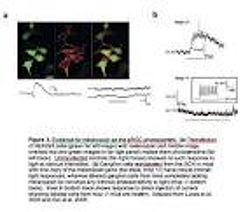
Рис.3. 3.[10] mice are healthy. Adapted from Lucas et al. 2003 and Qui et al. 2005.
- Рис. 3. Свидетельство меланопсина — фотопигмента фоторецептора ipRGC. Где:
(a)Трансфекция клеток HEK293 (зеленое крайне левое изображение) с melanopsin (красное среднее изображение, сверхположенные красные и зеленые изображения в далекой правильной группе) делает их фоточувствительными (крайне левый след). Средства управления Untransfected (далекие правильные следы) не показали такого ответа, чтобы осветить в различном intensities.
(b) клетки нервного узла retrolabeled от SCN в мышах с одной копией melanopsin гена (главный след, швабра +/-) имеют здравые свойственные легкие ответы, тогда как помеченные клетки нервного узла от мышей, полностью испытывающих недостаток melanopsin, не показывают никакой свойственной фоточувствительности, чтобы осветить (швабра-/-следоснования). Вставка в ответе показов следа основания, чтобы направить инъекцию потока, показывая помеченные клетки от швабры-/-здоровой мыши. Приспособленная Лукасом и др. 2003 и Qui и др. 2005.
Первые исследования клеток ipRGCs настоятельно определили меланопсин как фотопигмент в этих клетках и их роль для группы синего света, поглощающего известный «flavoproteins», поскольку cryptochromes не мог быть первоначально исключен (Berson 2007). Функция Cryptochromes как циркадные фотопигменты у беспозвоночных и одобрил их функционирование как циркадные фотопигменты, относящиеся к млекопитающим(Kavakli и 2002 Sancar; Ван Джелдер, Gibler и др. 2002). В настоящее время имеющиеся подавляющие свидетельства о том, что melanopsin является фотопигментом в ipRGCs, и, таким образом, истинный циркадный фотопигмент. Когда melanopsin ген удален посредством трансгенных методов в мышах (мыши нокаута), относящиеся к сетчатке глаза ячейки нервного узла, помеченные от SCN, которые не могут сигнализировать свет (Лукас, Hattar и др. 2003). Кроме того, животные с удалённым melanopsin-геном показывают дефициты в многократных визуальных отражениях, типа сжатия зрачка и фотозахвата (Панда, Sato и др. 2002; Рубин, Brennan и др. 2002; Лукас, Hattar и др. 2003; Панда, Provencio и др. 2003). Как дальнейший тест, Турция и. ал использовал множество мультиэлектродов, которые являются множеством плоских электродов, которое позволяет делать запись extracellularly от множеств относящихся к сетчатке глаза клеток нервного узла сразу, изолировать легкие ответы от многократных тестов на фоторецептоах ipRGCs. Сетчатки с клетками melanopsin мышей (нокаута) не показали никаких свойственных фотоответов от клеток нервного узла (Турция, Zhang и др. 2005). Несмотря на эти свидетельства, все еще были сомнения, споры относительно способности melanopsin функционировать в качестве истинного фотопигмента. Однако, они были твердо развеяны рядом изящных, тонких экспериментов, посредством выражения «ген-melanopsinа» в разнообразных типах клеток, которые становятся обычно нечувствительными к свету. Когда было сделано, чтобы выделить melanopsin ген, эти клетки оказались в состоянии сильно ответить на свет, указывая способность melanopsin функционировать как bonafide фотопигмент (см. Иллюстрацию 3) (Melyan, Tarttelin и др. 2005; Панда, Nayak и др. 2005; Qiu, Kumbalasiri и др. 2005).
Свет, зрение и биологические часы[править]
Свет перезагружает биологические часы в соответствии с кривой ответа фазы («СТРОИТЕЛЬСТВО ИЗ СБОРНОГО ЖЕЛЕЗОБЕТОНА») (PRC)en:Phase_response_curve. В зависимости от выбора времени, свет может продвинуть или задержать циркадный ритм. И «СТРОИТЕЛЬСТВО ИЗ СБОРНОГО ЖЕЛЕЗОБЕТОНА» и необходимая иллюминация изменяются от и до разновидностей, и более низкие легкие уровни освещения обязаны перезагружать часы у ночных грызунах, чем в людях.
Освещение уровней, которые затрагивают циркадный ритм в людях, выше чем уровни, обычно используемые в искусственном освещении в домах. Согласно некоторым исследователям[11]интенсивность освещения, которая взволновала циркадную систему, должна достигнуть до 1000 люксов, падающих на сетчатку.
В дополнении, чтобы осветить интенсивно, длина волны (или цвет) света — важный фактор в захвате биологических часов. Меланопсин — наиболее эффективно возбуждается при свете от синей части спектра согласно некоторым исследователям в диапазоне 420–440 нанометров[12], в то время как другие сообщили диапазоне 470-485 нанометров. Эти синие длины волн присутствуют в фактически всех источниках света, поэтому их устранение требует специальных огней или фильтров, которые кажутся янтарными.
Думается, что подбор света может иметь эффект определения циркадного ритма;[13] легкое освещение сверху, напоминая изображение яркого неба, имеет больший эффект, чем свет, входящий в наши глаза снизу, в виде отражённого.
Согласно исследованию 2010, законченному Исследовательским центром Освещения, дневной свет имеет прямой эффект на циркадные ритмы и, следовательно, на работу и благосостояние. Исследование показало, что, например, студенты, которые испытывают разрушение освещения схем утром, как правило, испытывают разрушение у спящих испытуемых представителей. Изменение у спящих может привести отрицательно в студенческой работе, на которую эот воздействует, что приводит к настороженности. Удаление циркадного света утром задерживает тусклое легкое начало мелатонина на 6 минут в день, в течение в общей сложности 30 минут в течение пяти дней.[14]
Это подтверждает, что меланопсин в фоторецепторах сечатки ipRGC в ганглиозном слое обладает низкой светочувствительностью, что биологически он приспособлен к встрече более сильных УФ лучей, являясь во-первых, фильтром и, самое главное, способен выполнять функции 24 часового циркадного ритма без участия палочек и колбочек, будучи связанным напрямую с мозгом, а также участвовать в подавлении сильных УФ лучей, автоматически, таким образом, участвовать в оппонентном отборе сильных цветовых лучей при цветном зрении и т.д.
В этой связи, последние исследования меланопсина даже на генетическом уровне (см. рис. 2,3) и др. исключают существующие сомнения, предположения о главной функции фотопигмента меланопсина — о его участии в формировании циркадных ритмов и в зрачковом рефлексе и др. — что этот фотопигмент фоторецепторов сетчатки ганглиозного слоя ipRGC — доказанный факт.[Замечание необходимое].
Шаблон:PBB Controls
Шаблон:Eye proteins
Шаблон:G protein-coupled receptors
—>
Примечания[править]
- ↑ https://webvision.med.utah.edu/Melanopsin.html
- ↑ https://c3012152.cdn.cloudfiles.rackspacecloud.com/110119eye.pdf
- ↑ Provencio I, Jiang G, De Grip W, Hayes W, Rollag M (1998). «Melanopsin: An opsin in melanophores, brain, and eye» (HTML: full text). Proc Natl Acad Sci U S A 95 (1): 340–5. DOI:10.1073/pnas.95.1.340. PMID 9419377.
- ↑ https://c3012152.cdn.cloudfiles.rackspacecloud.com/110119eye.pdf
- ↑ https://sciencenow.sciencemag.org/cgi/content/full/2010/111/1?etoc
- ↑ https://sciencenow.sciencemag.org/cgi/content/full/2010/111/1?etoc ref>[Ecker JL, Dumitrescu ON, Wong KY, Alam NM, Chen SK, LeGates T, Renna JM, Prusky GT, Berson DM, Hattar S (2010) Melanopsin-expressing retinal ganglion-cell photoreceptors: cellular diversity and role in pattern vision. Neuron 67:49–60]
- ↑ Provencio I, Jiang G, De Grip WJ, Hayes WP, Rollag MD (January 1998). «Melanopsin: An opsin in melanophores, brain, and eye». Proc. Natl. Acad. Sci. U.S.A. 95 (1): 340–5. DOI:10.1073/pnas.95.1.340. PMID 9419377.
- ↑ Roecklein, Kathryn (2009-04). «A missense variant (P10L) of the melanopsin (OPN4) gene in seasonal affective disorder.». Journal of Affective Disorders. https://www.ncbi.nlm.nih.gov/pubmed/18804284. Retrieved 2011-01-11.
- ↑ https://webvision.med.utah.edu/Melanopsin.html
- ↑ https://webvision.med.utah.edu/Melanopsin.html
- ↑ Semjonova, Milena (2003). «Healthy Lighting, from a lighting designer’s perspective». Milena Lighting Design. https://www.enlighter.org/images/2009/01/healthyLighting.pdf.
- ↑ Newman, L.A.; Walker, M.T.; Brown, R.L.; Cronin, T.W.; Robinson, P.R. (November 2003). «Melanopsin forms a functional short-wavelength photopigment». Biochemistry 42 (44): 12734–8. doi:10.1021/bi035418z. PMID 14596587.
- ↑ Semjonova, Milena (2003). «Healthy Lighting, from a lighting designer’s perspective». Milena Lighting Design. https://www.enlighter.org/images/2009/01/healthyLighting.pdf.
- ↑ Figueiro, M.G.; Rea, M.S. (February 2010). «Lack of short-wavelength light during the school day delays dim light melatonin onset (DLMO) in middle school students». Neuro Endocrinology Letters 31 (1): 4. PMID 20150866.
Источник

