
Глии в составе сетчатки
Сетчатка состоит из наружного пигментного и внутреннего светочувствительного слоёв. Внутренний её слой имеет зрительную (заднюю) и слепую (переднюю) части. На задней поверхности сетчатки находится слепое пятно — место выхода зрительного нерва. Несколько латеральнее от него располагается желтое пятно — участок наилучшего зрения. Сетчатка представляет цепь нейронов, между которыми находятся глиоциты и сосуды (рис. 23).
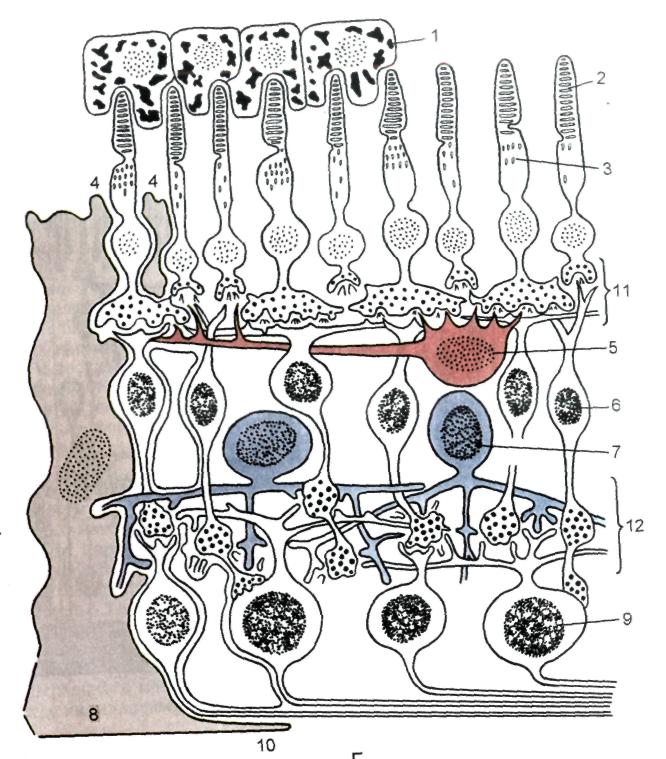
Рис. 23. Схема строения сетчатки. 1 – пигментоциты; 2 – палочки; 3 – колбочки; 4 – зона расположения наружной пограничной (глиальной) мембраны; 5 – гоРИзонтальные клетки; 6 – биполярные (вертикальные) клетки; 7 – амакринные клетки; 8 – глиоцит (мюллеровы волокна); 9 – ганглиозные клетки; 10 – зона расположения внутренней пограничной (глиальной) мембраны; 11 и 12 – синапсы.
В состав нейронов сетчатки входят светочувствительные (фоторецепторные), биполярные (ассоциативные) и ганглиозные нейроны, которые, располагаясь радиально, связаны между собой синапсами. Кроме этого, есть ещё два вида нейронов, обеспечивающих связь по горизонтали: горизонтальные клетки (соединяют фоторецепторные и биполярные нейроны) и амакринные клетки (соединяют биполярные и ганглиозные нейроны).
Фоторецепторные нейроны — типичные биполярные клетки. Их периферические отростки – дендриты имеют вид палочек и колбочек и образуют одноимённый слой. Центральные отростки – аксоны — образуют синаптические связи с биполярными и горизонтальными клетками.
И палочковые, и колбочковые отростки состоят из наружного и внутреннего сегментов, связанных ресничкой. Наружные сегменты располагаются между отростками клеток пигментного эпителия.
ПалочковИднЫе нейроны располагаются в периферических отделах сетчатки, воспринимают световые сигналы низкой интенсивности (сумеречное зрение) и отвечают за черно-белое зрение. Они имеют узкие, вытянутые периферические отростки (палочки), наружный сегмент которых имеет цилиндрическую форму и содержит стопкообразно расположенные мембранные диски (уплощощенные мешочки). В дисках находится зрительный пигмент родопсин. Под влиянием света родопсин расщепляется на составляющие: белок опсин и альдегид витамина А. Это влечёт гиперполяризацию рецепторов и изменение ионной проницаемости мембран, вследствие чего возникает биоэлектрический потенциал. В темноте осуществляется восстановление родопсина.
В проксимальных участках наружных сегментов происходит постоянное обновление дисков и их смещение в дистальные, где по мере старения они фагоцитируются пигментным эпителием. При недостаточном поступлении в организм витамина А обновление дисков нарушается, а при его отсутствии они разрушаются («куриная слепота»).
Внутренний сегмент палочек содержит удлиненные митохондрии, центриоли, элементы АЭС и ГЭС, комплекс Гольджи. Он обеспечивает наружный сегмент энергией и веществами, необходимыми для фоторецепции. Ядро палочковых фоторецепторов мелкое, округлое, окружено тонким ободком цитоплазмы. Аксоны заканчиваются шаровидными утолщениями.
КолбочковИдныЕ нейроны располагаются в центральных отделах сетчатки и особенно многочисленны в центральной ямке желтого пятна, реагируют на свет высокой интенсивности, обеспечивают дневное и цветовое зрение. Наружные сегменты колбочек имеют коническую форму. Они более короткие и широкие, чем у палочковых клеток. Мембранные диски в них образованы складками плазмолеммы и не отделены от неё. В мембране дисков содержится зрительный пигмент йодопсин, который в функционально различных типах колбочек разлагается под действием красного, зеленого или синего света. В колбочках, в отличие от палочек, не происходит постоянного перемещения дисков и их фагоцитоза пигментным эпителием. Строение внутреннего сегмента колбочек сходно с таковым у палочек. Ядра — крупнее и светлее, чем у палочковых клеток. Аксоны заканчиваются в наружном сетчатом слое расширением треугольной формы.
Биполярные (ассоциативные) нейроны своими дендритами образуют синапсы с аксонами фоторецепторных клеток, а аксонами — с дендритами ганглиозных и амакринных клеток. Подразделяются на несколько типов.
ГанглиоЗНые нейроны – самые крупные мультиполярные клетки с эксцентрично расположенным ядром и крупным ядрышком. В цитоплазме содержатся хорошо развитые органеллы. Дендриты образуют синапсы с аксонами биполярных клеток и отростками амакринных клеток, а аксоны образуют зрительный нерв.
Конвергенция (схождение) нервных импульсов в сетчатке обеспечивается характером связей ее нейронов и свойственна всем отделам, за исключением центральной ямки. Несколько палочковых клеток образуют синапсы на одной биполярной, а несколько биполярных контактируют с одной ганлионарной. Общий показатель конвергенции в сетчатке равен 105:1.
Горизонтальные нейроны — ассоциативные мультиполярные клетки; их дендриты и аксон синаптически связаны с аксонами палочковых и колбочковых клеток, а также с дендритами биполярных нейронов.
Амакринные клетки — ассоциативные нейроны, у которых, как правило, аксоны не выявляются, но некоторые из них содержат длинный аксоноподобный отросток. Дендриты образуют связи с аксонами биполярных клеток.
Пигментный эпителий сетчатки располагается на границе с сосудистой оболочкой и своими отростками проникает в фотосенсорный слой. Он образован кубическими (на периферии сетчатки) или призматическими (в ее центре) клетками с базально расположенными ядрами. В цитоплазме хорошо представлены органеллы общего значения и, особенно, лизосомы. Содержат большое количество гранул меланина. Базальная часть плазмолеммы имеет выраженную складчатость — признак активного ионного транспорта. Латеральные поверхности связаны десмосомами. Апикальная поверхность содержит микроворсинки и длинные ветвящиеся отростки, которые располагаются между палочками и колбочками фоторецепторных нейронов.
В функциональном плане пигментный эпителий осуществляет трофику фоторецепторных нейронов, транспорт к их периферическим отросткам витамина А, и др. веществ, способствующих регенерации; фагоцитоз и переваривание кончиков наружных сегментов палочковых фоторецепторов; обеспечение избирательной диффузии веществ из сосудисто-капиллярного слоя сосудистой оболочки; регуляцию поступления светового потока к рецепторам и предотвращение избыточной их засветки, благодаря синтезу и перемещению меланина в отростки на свету. В темноте гранулы меланина перемещаются из отростков в тело клеток.
Нейроглия сетчатки представлена радиальными глиоцитами {мюллеровыми клетками), астроцитами и микроглией.
Мюллеровы клетки пронизывают почти всю толщину сетчатки. Своими основаниями они на границе со стекловидным телом формируют внутреннюю глиальную пограничную мембрану, а апикальными отростками образуют у основания палочек и колбочек наружную глиальную пограничную мембрану. Выполняют поддерживающую и трофическую функции. Вместе с астроцитами они образуют гемато-ретинальный барьер.
Микроглиоциты немногочисленны, располагаются во всех слоях сетчатки, осуществляют фагоцитоз.
Слои сетчатки:
Периферические отростки фотосенсорных нейронов образуют слой палочек и колбочек, ядросодержащие участки клеток — наружный, внутренний ядерные и ганглиозный слои, а области синаптических связей нейронов – наружный и внутренний сетчатые слои. Аксоны ганглиозных нейронов формируют слой нервных волокон, образующих зрительный нерв.
Кроме того, слой палочек и колбочек отделён от наружного ядерного слоя внутренней глиальной пограничной мембраной, а на границе слоя нервных волокон и стекловидного тела располагается наружная глиальная пограничная мембрана.
Кровоснабжение сетчатки осуществляется ветвями центральной артерии сетчатки. Проникая вместе со зрительным нервом, она в области сосочка разделяется на радиально расположенные ветви, которые сначала располагаются между стекловидным телом и внутренней глиальной пограничной мембраной, а затем заходят в слои сетчатки, образуя капиллярное сплетение, доходящее до внутреннего ядерного слоя. Трофика расположенных кнаружи от него слоёв сетчатки осуществляется за счёт сосудисто-капиллярного слоя сосудистой оболочки диффузно через слой пигментного эпителия.
Источник
Глиальная система сетчатой оболочки выполняет те же функции, что глия центральной нервной системы. В сетчатке различают четыре типа клеток: мюллеровская клетка, астроциты, олигодендроциты и микроглия [39, 496, 799, 800, 1008]. Некоторые авторы выделяют еще один тип глии — специализированный астроцит, который располагается только вблизи кровеносных сосудов (периваскулярная глия Лисса).
Астроглия(рис. 3.6.41). Астроциты возникают в эмбриональном периоде из клеток нев-рального гребня, проникая в сетчатку по ходу зрительного нерва [189, 1043]. Различают «фиброзный» и «протоплазматический» астроциты [492—495, 1185]. Типичной особенностью аст-роцитов центральной нервной системы, в том числе сетчатки, являются длинные маловетвя-щиеся отростки, часть которых примыкает к стенкам небольших кровеносных сосудов. Тело клетки и ядро имеют овальную и полигональную форму и слабо окрашены. В ядре содержится небольшое количество хроматина. Ядрышко, как правило, обнаружить не удается. Цитоплазма астроцитов насыщена микрофила-ментами (10 нм в диаметре). Хорошо развит эндоплазматический ретикулум. Видны гранулы гликогена, длинные митохондрии, центриоли и реснички [1008]. Фибриллы могут объединяться в пучки различной толщины и длины. Иммуно-гистохимически как в цитоплазме клеток, так и в их отростках выявлен маркерный белок — фибриллярный кислый белок глии [752].
Фиброзные астроциты содержат мало митохондрий и больше микрофиламентов, чем про-топлазматические астроциты.
Сетчатка
i ^ЁШШ!^л,^&<Ш^&ш^~ Si *»
Рис. 3.6.41. Особенности распределения астроцитов по периферии (а) и в центральных (б) участках слоя нервных волокон сетчатой оболочки (по Schnitzer, 1988)
Отростки протоплазматических астроцитов более короткие и толстые. Простираются они во внутреннем плексиформном слое. Их ядра различного размера и содержат грубые зерна гетерохроматина. Как тела клеток, так и их отростки располагаются только в слое нервных волокон сетчатки. Причем морфология клеток изменяется в различных участках сетчатки. Вблизи диска зрительного нерва их отростки исключительно длинные, а по периферии клетки принимают звездчатую форму с одинаковой длины более короткими отростками. Астроциты отсутствуют в области желтого пятна и зубчатой линии. Вообще, число астроцитов коррелирует с толщиной слоя нервных волокон сетчатки, в котором разветвляются их отростки [166].
Астроциты охватывают, особенно при проникновении в склеральный канал, аксоны ганг-лиозных клеток, формируя вокруг них футляр (рис. 3.6.41, 3.6.42).
Особенностью астроцитов является и то, что они контактируют с кровеносными сосудами, образуя при этом щелевые контакты, расположенные на их ножках. Между собой они соединяются при помощи щелевых контактов и зон слипания. Предполагают, что это взаимодействие обеспечивает функционирование гема-тоэнцефалического барьера.
Рис. 3.6.42. Объемное схематическое изображение взаимоотношения астроцитов с пучками аксонов ганглиоз-ных клеток и кровеносными сосудами в слое нервных волокон сетчатки:
/ — астроциты; 2 — аксоны ганглиозных клеток; 3 — кровеносные сосуды
Подобно мюллеровским клеткам, астроциты обеспечивают нейроны глюкозой и участвуют в поддержании ионного состава межклеточной жидкости. Кроме того, астроциты поддерживают нормальный уровень метаболизма нейроме-диаторов.
Одной из основных функций астроцитов является защитная функция. При повреждении ткани сетчатки астроциты подвергаются гипертрофии и размножаются, образуя глиальный рубец [799, 800]. Процесс регуляции пролифе-ративной активности астроцитов в норме и при патологичесих состояниях (глаукома) как сетчатки, так и зрительного нерва находится под контролем эндотелина-1.
Олигодендроциты.Классические формы олигодендроглиальных клеток свойственны зрительному нерву. В сетчатке большинства позвоночных животных клетки, напоминающие олигодендроциты, располагаются в слое ганглиозных клеток [37, 38, 19, 882]. В сетчатке человека этот тип клеток рядом исследователей не выделяется [154]. Тем не менее некоторые исследователи на основании общности функции мюл-леровских клеток и клеток олигодендрогии считают эти клетки близкими по происхождению.
Олигодендроциты позвоночных являются самыми мелкими клетками ганглиозного слоя [39].
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
Их форма округлая или овальная. Ядрышко небольшое и расположено в центре ядра. Для клеток этого типа характерно расположение группами по 2—3 клетки в непосредственной близости от крупных нейронов. Именно поэтому их количество существенно возрастает при увеличении концентрации нейронов. Среди клеток редко встречаются митозы.
Как и остальные глиальные элементы сетчатой оболочки, олигодендроциты образуют единую функционально-метаболическую систему с нейронами сетчатки [25, 26].
По всей видимости, олигодендроциты, расположенные в слое ганглиозных клеток, не способны к миелогенезу. Появляется эта способность лишь в области диска зрительного нерва при формировании миелиновой оболочки аксонов ганглиозных клеток.
Микроглия(рис. 3.6.43). Микроглия складывается из маленьких клеток (до 30 мкм), имеющих мезодермальное происхождение [39, 189, 1008]. В ганглиозном слое у всех позвоночных микроглиоциты часто являются сателлитами нейронов, а свои цитоплазматические отростки посылают к капиллярам, оплетая их.
шшжш
Рис. 3.6.43. Локализация и особенности строения мик-роглиальных клеток сетчатки:
а — локализация микроглиальных клеток (импрегнация по Гольд-жи); б — лектин-окрашенная клетка микроглии (по Chan-Ling,
1994)
Различают два типа микроглиальных клеток. Один тип клеток мигрирует в сетчатку на наиболее ранних этапах эмбрионального развития вместе с мезенхимой зрительного нерва. Второй тип клеток поступает в сетчатку из кровеносного русла (моноциты) или исходят из перицитов кровеносных сосудов [99, 125, 189].
Цитоплазма микроглиальных клеток напоминает цитоплазму астроцитов, но при этом в ней меньше гранул гликогена и меньше микрофила-ментов. Цитоплазма скудная, а ядро светлое. Отличительной особенностью микроглиальных клеток является насыщение цитоплазмы длинными профилями шероховатой эндоплазмати-ческой сети, наличием небольшого количества микротрубочек. В цитоплазме можно также обнаружить многочисленные лизосомы и липо-фусциновые гранулы.
Клетки микроглии распределены равномерно во всей толще сетчатой оболочки, но неравномерно по площади сетчатки. Необходимо подчеркнуть, что микроглиальные клетки являются единственным глиальным элементом слоя Хен-ле в области центральной ямки.
Функции микроглии сетчатки до сих пор полностью не выяснены. По происхождению, форме, топографии и по аналогии с гистиоцитами центральной нервной системы их можно отнести к фагоцитирующим и переваривающим клеткам [39]. В отличие от макроглиальных клеток микроглия не участвует в процессах репарации. После травмы они размножаются и начинают напоминать гистиоциты [709]. При этом они фагоцитируют продукты распада клеточных элементов сетчатки. Как и в головном мозге, микроглиальные клетки способны к амебоидному передвижению (трансформируются в макрофаги) [ИЗО, 1034]. Таким образом, основной функцией микроглии является защитная функция. Это особенно четко проявляется при различных патологических состояниях как сетчатой оболочки, так и увеального тракта [1185].
Клетки Мюллера(рис. 3.6.44, 3.6.45). Мюл-леровские клетки являются самыми крупными клетками сетчатой оболочки. Распространяются они от наружной пограничной мембраны до внутренней пограничной мембраны [39]. Средняя плотность мюллеровских клеток примерно равна 8000—13 000 клеток в мм2[264].
В эмбриональном периоде мюллеровские клетки возникают из внутреннего слоя зрительного бокала в два этапа [1116]. На самых ранних этапах нейроэпителиальные клетки края глазного бокала, смежные с клетками будущего пигментного эпителия сетчатки, образуют первичные нейроны (колбочки, горизонтальные клетки и ганглиозные клетки). Второй этап развития нейроэпителиальных клеток приводит к образованию палочек, биполярных, амакри-новых клеток, а также мюллеровских клеток [885]. Все развивающиеся нейроны и мюллеровские клетки мигрируют к месту своего постоян-
Источник
Нейроны
сетчатки синтезируют ацетилхолин,
дофамин, L-глутаминовую кислоту (например,
между фоторецепторными нейронами и
биполярными клетками), глицин,
гама-аминомасляную кислоту. Некоторые
нейроны содержат серотонин, его аналоги
(индоламины) и нейропептиды.
Горизонтальные
клетки.
Перикарионы расположены в наружной
части внутреннего ядерного слоя, а
отростки входят в область синапсов
между фоторецепторными и биполярными
клетками. Горизонтальные клетки получают
информацию от колбочек и передают её
также колбочкам. Соседние горизонтальные
клетки связаны между собой щелевыми
контактами.
Амакринные
клетки.
Их перикарионы находятся во внутренней
части внутреннего ядерного слоя в
области синапсов между биполярными и
ганглиозными клетками.
Биполярные
клетки реагируют на контрастность
изображения. Некоторые биполяры сильнее
реагируют на цветной, нежели на чёрно-белый
контраст. Одни получают информацию
преимущественно от палочек, другие —
от колбочек.
Ганглиозные
клетки —
крупные мультиполярные нейроны многих
разновидностей. Их аксоны образуют
зрительный нерв. Ганглиозные клетки
реагируют на множество свойств зрительного
объекта (например, на светлые и тёмные
объекты, однородность освещения, цвет
объекта, его ориентацию).
Сетчатка
Глия сетчатки
Кроме
нейронов, сетчатка содержит крупные
клетки радиальной глии. Их ядра расположены
на уровне центральной части внутреннего
ядерного слоя. Наружные отростки
заканчиваются микроворсинками, образуя
наружный пограничный слой. Внутренние
отростки имеют расширение (ножку) во
внутреннем пограничном слое на границе
со стекловидным телом. Глиальные клетки
играют важную роль в регуляции ионного
гомеостаза сетчатки. В частности, они
снижают концентрацию K+ во внеклеточном
пространстве, где концентрация этих
ионов при световом раздражении резко
увеличивается. Плазматическая мембрана
радиальной глии в области ножки
характеризуется высокой проницаемостью
для ионов K+, выходящих из клетки. Клетка
радиальной глии захватывает K+ из наружных
слоёв сетчатки и направляет поток этих
ионов через свою ножку в жидкость
стекловидного
тела.
І.
Склера;
ІІ.
Собственно сосудистая оболочка; ІІІ.
Сетчатка.
Механизм фотовосприятия
В
состав дисков фоторецепторных клеток
входят зрительные пигменты, в том числе
родопсин палочек.
Родопсин
состоит из белковой части (опсин) и
хромофора — 11-цис-ретиналя, под
действием фотонов переходящего в
транс-ретиналь. Мутации генов, кодирующих
синтез опсинов, приводят к развитию
пигментного ретинита и ночной (куриной)
слепоты. Описано около 40 мутаций генов
опсинов.
Нас
всех учили в школе, в институтах, в
научных и популярных статьях и книгах,
что глаз человека устроен подобно
фотоаппарату. «Объектив» глаза — хрусталик
проектирует изображение на чувствительные
элементы сетчатки — торцы палочек и
колбочек, которые образуют
«экран-фотопластинку». Сигналы от них
не исследованными до конца путями
попадают в мозг по глазному нерву. Он
реально является жгутом многих нервных
волокон, число которых на порядки меньше
числа палочек и колбочек. Удавалось
даже найти в областях мозга, ответственных
за зрение, что-то похожее на нерезкую
проекцию изображения, попадающего в
глаз. Однако откройте физический,
биологический, медицинский учебник.
Там обычно приводится сечение сетчатки
глаза. Слои клеток, получившие свои
названия по произволу их открывателей.
Вопреки всему, что объясняли нам в школе,
она направлена не на торцы палочек и
колбочек, а через вспомогательные
нервные клетки (нейроны) в сетчатке
глаза на обратную
сторону
палочек и колбочек! Торцы палочек и
колбочек не могут ничего «видеть», так
как они упёрты в непрозрачный тёмный
пигментный слой. В аналогиях с техническими
устройствами свет на сетчатку глаза
падает не на «фотодиоды», а на
«технологическую плату» сзади них, на
которой они «распаяны». Об этом учебники
и научные статьи напрочь стыдливо
молчат. Нонсенс!
Глаз
вместе с мозгом — «компьютер», обрабатывающий
спектры пространственных частот и их
функции корреляции, а не аналог
«фотоаппарата».
Адаптация
зрения к условиям освещения. У
разных животных разные наборы палочек
и колбочек. У строго дневных ящериц
только колбочковая сетчатка. У животных,
активных в с умерки, в сетчатке преобладают
палочки. У животных, активных и днем, и
в сумерки, сетчатка содержит и палочки,
и колбочки. После заката солнца у таких
животных идет «перестройка» сетчатки
с колбочкового зрения на палочковое —
так называемая темновая адаптация. Так,
человек в сумерки перестает различать
цвета («ночью все кошки серы»): красные
цветы мака становятся черными, а
сине-фиолетовые — очень светлыми. Это
происходит потому, что максимум
спектральной чувствительности у палочек
сдвинут относительно колбочек в голубой
конец спектра. Это явление носит название
сдвига Пуркинье (по имени чешского
естествоиспытателя Я. Пуркине. В сетчатке
млекопитающих темновая адаптация идет
за счет перестройки нервных связей
внутри сетчатки, у рыб — за счет движения
рецепторов в сетчатке: при большой
яркости освещения палочки выдвигаются
и «прячутся» от света в отростках клеток
пигментного эпителия, а в эти отростки
заходят гранулы темного экранирующего
пигмента меланина. В сумерки, напротив,
колбочки уползают от света, а палочки
приближаются. Это так называемая
ретиномоторная реакция. Колбочки низших
позвоночных, содержащие разные зрительные
пигменты, имеют и разное строение. Они
располагаются в сетчатке регулярно,
образуя разные мозаичные картины,
характерные для данного вида животных.
У рептилий (кроме змей и гекконов) и
дневных птиц в колбочках между наружным
и внутренним сегментами, т. е. на пути
света, находится жировая капля, окрашенная
каротиноидными пигментами, из-за чего
сетчатка ящерицы или черепахи под
микроскопом выглядит как ткань в красный,
оранжевый и желтый горошек. Эти
внутриколбочковые фильтры изменяют
реальную спектральную чувствительность
колбочки. Возможно, они служат линзами,
фокусирующими свет на наружном сегменте,
или предохраняют наружные сегменты от
повреждающего действия ультрафиолета.
У млекопитающих все колбочки одинаковой
формы. В сетчатке человека и обезьян
колбочки, содержащие разные зрительные
пигменты, расположены хаотически.
Разными наборами рецепторов определяются
свойства зрения животного, в частности
его способность воспринимать цвета
Источник

