
Ганглиозные клетки сетчатки связанные с рецептивным полем возбуждаются при
Оглавление темы «Рецепторный потенциал палочек и колбочек. Рецептивные поля клеток сетчатки. Проводящие пути и центры зрительной системы. Зрительное восприятие.»:
1. Рецепторный потенциал палочек и колбочек. Ток ионов через мембрану фоторецептора в темноте и на свету.
2. Адаптация фоторецепторов к изменениям освещенности. Световая адаптация. Десенситизация. Темновая адаптация.
3. Рецептивные поля клеток сетчатки. Прямой путь передачи сигналов от фоторецепторов к ганглиозной клетке. Непрямой путь передачи сигналов.
4. Рецептивные поля с оn-центрами и off-центрами. On-нейроны. Off-нейроны. Ганглиозная клетка on-типа. Ганглиозная клетка off-типа.
5. Рецептивные поля цветового восприятия. Восприятие цвета. Первичные цвета. Монохромазия. Дихромазия. Трихромазия.
6. М- и Р-типы ганглиозных клеток сетчатки. Магноцеллюлярные (М-клетки) клетки. Парвоцеллюлярные (Р-клетки) ганглиозные клетки сетчатки.
7. Проводящие пути и центры зрительной системы. Зрительный нерв. Зрительные тракты. Глазодвигательный рефлекс.
8. Латеральное коленчатое тело. Функциональная организация латерального коленчатого тела. Рецептивные поля латерального коленчатого тела.
9. Переработка зрительной сенсорной информации в коре. Проекционная зрительная кора. Световая грань. Комплексные нейроны. Двойные противоцветные клетки.
10. Зрительное восприятие. Магноцеллюлярный путь. Парвоцеллюлярный путь. Восприятие формы, цвета.
Рецептивные поля с оn-центрами и off-центрами. On-нейроны. Off-нейроны. Ганглиозная клетка on-типа. Ганглиозная клетка off-типа.
В сетчатке человека имеются два типа ганглиозных клеток, отличающихся реакцией на точечные световые стимулы, воздействовавшие на центр или периферию их рецептивного поля (рис. 17.9). Примерно половина ганглиозных клеток возбуждается действием света на центр рецептивного поля и тормозится при действии светового стимула на периферию рецептивного поля. Такие клетки принято называть оn-нейронами. Другая половина ганглиозных клеток возбуждается действием светового раздражителя на периферию рецептивного поля и тормозится в ответ на световую стимуляцию центра рецептивного поля — они получили название off-нейронов.
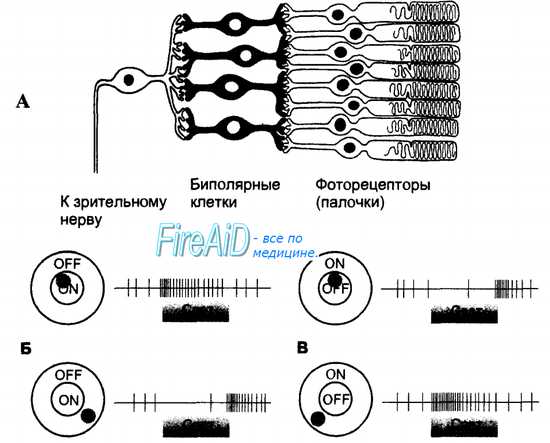
Рис. 17.9. Рецептивные поля ганглиозных клеток сетчатки с on- и off-центрами.
A. Рецептивное поле ганглиозной клетки образуют все фоторецепторы и биполярные клетки, имеющие с нею синаптические контакты. Ганглиозные клетки постоянно генерируют потенциалы действия, частота возникновения которых зависит от активности фоторецепторов и биполярных клеток, входящих в ее рецептивное поле.
Б. Ганглиозная клетка on-типа увеличивает частоту электрических разрядов в ответ на световое раздражение центра рецептивного поля и снижает свою электрическую активность при действии светового раздражителя на периферию рецептивного поля.
B. Ганглиозная клетка off-типа тормозится при действии света на центр ее рецептивного поля и увеличивает частоту нервных импульсов в ответ на раздражение периферии рецептивного поля.
Рецептивные поля ганглиозных клеток обоих типов в сетчатке представлены поровну, чередуясь друг с другом. Оба типа клеток очень слабо отвечают на равномерную диффузную засветку всего рецептивного поля, а наиболее сильным раздражителем для них является световой контраст, т. е. различная интенсивность засветки центра и периферии. Именно контрастирование деталей изображения дает необходимую информацию для зрительного восприятия в целом, тогда как абсолютная интенсивность отраженного от наблюдаемого объекта света не столь важна. Восприятие граней, т. е. восприятие контраста между соседними поверхностями с разной освещенностью, является наиболее информативным признаком изображения, определяющим протяженность и позиции разных объектов.
— Также рекомендуем «Рецептивные поля цветового восприятия. Восприятие цвета. Первичные цвета. Монохромазия. Дихромазия. Трихромазия.»
Источник
Ганглионарная (ганглиозная) клетка — нервная клетка (нейрон) сетчатки глаза, способная генерировать нервные импульсы в отличие от других типов нейронов сетчатки (биполярных, горизонтальных, амакриновых). В их цитоплазме хорошо выражено базофильное вещество. Ганглионарные клетки граничат со стекловидным телом глаза и образуют слой сетчатки, который первым получает свет. Их аксоны по поверхности сетчатки направляются к слепому пятну (пятно Мариотта), собираются в зрительный нерв и направляются в мозг. Аксоны ганглионарных клеток не миелинизированы при прохождении сетчатки, чтобы не препятствовать прохождению света. Далее они покрыты миелиновой оболочкой.
Ганглионарные клетки завершают «трёхнейронную рецепторно-проводящую систему сетчатки»: фоторецептор — биполярный нейрон — ганглионарная клетка.
Функции ганглионарных клеток[править | править код]
Клетки сетчатки связаны между собой сложной сетью возбуждающих, подавляющих и двунаправленных сигнальных связей. Они собирают информацию от всех слоев сетчатки как по вертикальным путям (фоторецепторы — биполяры — ганглионарные клетки), так и по латеральным путям (фоторецепторы — горизонтальные клетки — биполяры — амакриновые клетки — ганглионарные клетки).
Рецептивные поля[править | править код]
На одну ганглионарную клетку приходится от одного до сотни биполярных нейронов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от нескольких до нескольких тысяч фоторецепторов. Каждая ганглионарная клетка суммирует сигналы от большого числа фоторецепторов, что повышает световую чувствительность, но уменьшает разрешение. Фоторецепторы, соединенные с одной ганглионарной клеткой, образуют её рецептивное поле. Рецептивные поля ганглионарных клеток подразделяются на простые и сложные. Простые поля — имеют концентрическую структуру, подобно полям биполярных нейронов. Сложные — разделяются на несколько различных функциональных зон. Рецептивные поля могут перестраиваться, адаптируясь к уровню освещённости и характеристикам световых стимулов. Через биполярные нейроны с одной ганглионарной клеткой может быть связано от единиц до десятков тысяч фоторецепторов (палочек и колбочек). В свою очередь, один фоторецептор через биполярные нейроны может быть связан с десятками ганглионарных клеток. В среднем на 100 фоторецепторных клеток приходится одна ганглионарная (т.е., их от 1.2 до 1.5 млн). Чем ближе к центральной ямке глаза, тем меньше фоторецепторов приходится на одну ганглионарную клетку. Поэтому у людей слабое периферическое зрение. В районе центральной ямки, наоборот, высокое разрешение, но менее высокая светочувствительность, поскольку здесь каждый фоторецептор (колбочка) соединен с одной биполярной (карликовой) клеткой, которая в свою очередь соединена лишь с одной ганглионарной.
Типы ганглионарных клеток[править | править код]
Существует всего 18 типов ганглионарных клеток сетчатки.
Большинство относится к трем типам:
1. Парвоганглионарные клетки — карликовые клетки (около 80% от числа всех ганглионарных клеток сетчатки), имеющие средний размер тела и маленькое дерево дендритов, входят в карликовый путь (чувствительный путь, ведущий от глаза к четверохолмию) и связаны с парвоцеллюлярными (мелкоклеточными) слоями латеральных коленчатых тел. С этими клетками связывают высокую остроту зрения и цветовое зрение.
2. Магноклетки — (около 10%) очень разнообразны (малые и большие зонтичные клетки): с большими телами и многочисленными укороченными ветвями, маленькими телами и большим разветвлением дендритов, которые проецируются в крупноклеточные слои латеральных коленчатых тел. Отвечают за востриятие движущихся объектов. Имеют большие рецепторные поля.
3. Кониоцеллюлярные клетки очень мелкие, составляют от 8 до 10% всех ганглионарных клеток сетчатки. Получают сигналы от среднего количества фоторецепторов. Имеют очень большие рецептивные поля. Всегда ON для колбочек синего цвета и OFF для красного и зеленого.
Выделяют ганглионарные клетки, связанные с палочковыми и колбочковыми нейронами, с on- и off-центрами, которые отвечают на световое раздражение деполяризацией или гиперполяризацией соответственно. Дендриты клеток с on-центром разветвляются в подуровне а (пигментном эпителии?), с off-центром в подуровне G (ганглионарном слое?) внутреннего сетчатого слоя. Цветовой канал связан с красным, зеленым и синим типом on/off-ганглионарных клеток. Если красный и зеленый тип ганглионарных on/off-клеток относится к карликовому пути, то синий тип не относится к последнему. On/off-ответы ганглионарных клеток определяются специальными контактами колбочковых биполяров и расположением ганглионарных клеток в соответствующем подуровне внутреннего сетчатого слоя
Светочувствительные ганглионарные клетки[править | править код]
В 1991 году были открыты особые светочувствительные ганглионарные клетки типа ipRGC (intrinsically photosensitive retinal ganglion cells), или mRGC (melanopsin-containing retinal ganglion cells). Они, в отличие от ранее известных ганглионарных клеток, содержат светочувствительный пигмент меланопсин, отличающийся от других фоточувствительных пигментов глаза: родопсина палочек и йодопсина колбочек. И этим они отличаются от других ганглионарных клеток, находящихся в сетчатке глаза, которые не умеют реагировать непосредственно на свет.
Эти светочувствительные ганглионарные клетки — новый, третий тип фоторецепторов сетчатки глаза, помимо известных уже в течение 200 лет палочек и колбочек. Они напрямую возбуждаются под действием света даже при блокировании «классических» фоторецепторов глаза — палочек и колбочек.
Нервные пути от этих ганглиозных (ганглионарных) клеток ведут порождённое в них светом возбуждение от сетчатки к гипоталамусу тремя разными путями, обеспечивая световое управление циркадными ритмами, а также по отдельному нервному пути обеспечивают реакцию сужения зрачка на свет.
Литература[править | править код]
- Ноздрачёв А. Д., Баженов Ю. И., Баранникова И. А., Батуев А. С. и др. Начала физиологии: Учебник для вузов / Под ред. акад. А. Д. Ноздрачёва. СПб.: Лань, 2001. 1088 с.
Ссылки[править | править код]
- Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System
- Photosensitive ganglion cells
- Vision beyond image formation: The role of melanopsin cells in regulating mammalian physiology
- Blind Mice Can «See» Thanks To Special Retinal Cells
- Фоторецепторы и фоторецепция
[1]
[2]
[3]
[4]
[5]
Источник
[править]
Материал из Википедии — свободной энциклопедии
Рецептивное поле (англ. receptive field) сенсорного нейрона — участок с рецепторами, которые при воздействии на них определённого стимула приводят к изменению возбуждения этого нейрона.
Концепция рецептивных полей может быть применима ко всей нервной системе. Если множество сенсорных рецепторов образуют синапсы c единственным нейроном, они совместно формируют рецептивное поле этого нейрона. Например, рецептивное поле ганглионарной (ганглиозной) клетки сетчатки глаза представлено фоторецепторными клетками (палочками или колбочками), а группа ганглионарных клеток в свою очередь создаёт рецептивное поле для одного из нейронов мозга. В итоге к одному нейрону более высокого синаптического уровня сходятся импульсы от многих фоторецепторов; и этот процесс называется конвергенцией.
[править] Слуховая система
В качестве рецептивных полей слуховой системы могут рассматриваться части слухового пространства (англ. auditory space) или диапазоны слуховых частот. Лишь немногие исследователи трактуют слуховые рецептивные поля как определённые участки сенсорного эпителия, например, группы волосковых клеток спирального органа улитки внутреннего уха млекопитающих.
[править] Соматосенсорная система
Рецептивные поля соматосенсорной системы — это участки кожи или внутренних органов. Размеры рецептивных полей неодинаковы и зависят от типа механорецепторов.
Большое рецептивное поле нейрона позволяет отслеживать изменения на
большей площади чувствительной поверхности, но обеспечивает меньшую
разрешающую способность ощущения. Таким образом, пальцы, которые должны осязать тонкие детали, имеют множество плотно расположенных (до 500 на 1 см3) механорецепторов с маленькими рецептивными полями (около 10 мм2), тогда как спина, бёдра и голени
имеют меньшее количество рецепторов, объединённых в большие рецептивные
поля. Как правило, в центральной части большого рецептивного поля
имеется одно «горячее пятно», стимуляция которого вызывает наиболее
интенсивный ответ.
Нейроны коры головного мозга, связанные с тактильными ощущениями, имеют рецептивные поля на коже,
размеры и расположение которых могут изменяться по мере накопления
человеком индивидуального опыта или вследствие повреждения сенсорных
(афферентных) нервных волокон. В основном эти нейроны имеют относительно большие рецептивные поля — гораздо большего размера, чем у нейронов спинномозгового узла (спинального ганглия). Тем не менее, благодаря особым механизмам возбуждения и торможения в рецептивных полях, улучшающим пространственное разрешение, эти кортикальные нейроны способны распознавать тонкие детали.
[править] Зрительная система
Рецептивные поля зрительной системы можно считать частями зрительного пространства (англ. visual space). Например, в качестве рецептивного поля одной фоторецепторной клетки можно рассматривать конус, охватывающий все возможные направления, с которых эта клетка способна воспринимать свет. Его вершина находится в центре хрусталика, а основание — в бесконечности зрительного пространства. Но традиционно зрительные рецептивные поля изображаются на плоскости — как круги, квадраты, прямоугольники… Подобные изображения по сути являются сечениями конуса, отвечающего рецептивному полю одной специфической клетки, плоскостью, в которой исследователь предъявлял конкретный визуальный стимул. Рецептивные поля бинокулярных нейронов первичной зрительной коры (или стриарной области — поле Бродмана 17, зрительная зона V1) не уходят в оптическую бесконечность, а ограничены определённым расстоянием от точки, в которую направлен взгляд — «точки фиксации глаз» (См. зона Панума — англ. Panum’s area).
Рецептивные поля нейронов зачастую определяются как области сетчатки,
освещение которых изменяет возбуждение конкретного нейрона. Для
ганглионарных (ганглиозных) клеток сетчатки эта область включает все
фоторецепторы — палочки или колбочки одного глаза, связанные с
конкретной ганглионарной клеткой посредством синаптических контактов с биполярными, горизонтальными и амакринными (амакриновыми) клетками. Для бинокулярных нейронов зрительной коры (англ. visual cortex)
рецептивные поля определяются как совокупность соответствующих областей
сетчаток правого и левого глаза. Эти области могут быть закартированы
по отдельности в каждой сетчатке (при закрывании другого глаза), но
полностью связь каждой из областей с возбуждением исследуемого нейрона
обнаруживается только в том случае, когда открыты оба глаза.
Хьюбел и Визель
(например, Hubel, 1963) развили теорию о том, что рецептивные поля
клеток каждого уровня зрительной системы формируются синаптическими
соединениями с клетками более низкого иерархического уровня этой
системы. В этом случае небольшие и просто устроенные рецептивные поля
могут комбинироваться, формируя обширные и сложные рецептивные поля.
Позднее нейробиологи
усовершенствовали эту относительно простую концепцию, допустив, что
нейроны низших уровней зрительной системы связаны обратными эфферентными
связями с нейронами более высоких уровней.
В настоящее время составлены карты рецептивных полей для клеток всех
уровней зрительной системы — от фоторецепторов и ганглионарных
(ганглиозных) клеток сетчатки — до нейронов латерального (наружного) коленчатого тела,
первичной и экстрастриарной зрительной коры. Исследования, основанные
лишь на ощущениях, не могут дать полной картины для понимания феномена
зрения, поэтому здесь, также как и при изучении мозга, должны
применяться электрофизиологические методы — тем более, что в
эмбриогенезе млекопитающих сетчатка возникает в процессе дальнейшей
дифференциации латеральных выпячиваний промежуточного мозга (так
называемых глазных пузырей).[1]
[править] Ганглионарные клетки сетчатки
Ганглионарные (ганглиозные) клетки сетчатки
с on — и off — центрами отвечают диаметрально противоположным образом на освещение центра и периферии рецептивного поля.
Сильный ответ соответствует высокочастотному возбуждению, слабый — низкочастотному, отсутствие ответа — отсутствию активности.
Распознавание границ изображения (краёв, углов) рецептивными полями сетчатки — грубая компьютерная аппроксимация.
Размеры рецептивных полей увеличиваются от центра сетчатки к её периферии.
Визуальная информация от двух типов клеток (с on- и off- центрами) показана красным и зелёным цветом, соответственно.
Каждая ганглионарная (ганглиозная) клетка или оптическое нервное волокно (англ. optic nerve fiber)
порождает рецептивное поле, расширяющееся по мере возрастания
интенсивности освещения. Если размер поля максимален, то свет на его
периферии интенсивнее, нежели в центре, отражая то, что некоторые
синаптические пути предпочтительнее других.
Организация рецептивных полей ганглиозных клеток, составленных из входов многих палочек или колбочек, позволяет обнаруживать контраст,
что используется для выявления краевых частей наблюдаемых объектов.
Каждое рецептивное поле подразделяется на две части: центральный диск — «центр» и концентрическое кольцо — «периферию»;
каждая из этих частей реагирует на свет противоположным образом. Так,
если освещение центра рецептивного поля увеличивает возбуждение
конкретной ганглиозной клетки с так называемым on-центром (см. далее), то воздействие света на периферию этого же поля оказывает тормозящее воздействие на эту ганглиозную клетку.
Существует два основных типа ганглиозных клеток: с «on-центром» и «off-центром». Клетка с on-центром возбуждается при освещении центра и тормозится при освещении периферии её рецептивного поля. Реакция на свет клетки с off-центром диаметрально противоположная. Кроме того, у млекопитающих имеются клетки промежуточного (on-off) типа, которым свойственна кратковременная реакция на освещение по on-типу и на затенение по off-типу.[2] Освещение центральной части рецептивного поля приводит к деполяризации и возрастанию возбуждения нейрона (например, ганглионарной клетки) с on-центром, освещение периферии рецептивного поля приводит к гиперполяризации
и торможению этого нейрона, а одновременная световая стимуляция и
центра, и периферии рецептивного поля вызывает слабую активацию
(вследствие суммации эффектов, связанных с реакциями центральной и
периферической частей рецептивного поля). Ганглионарная клетка (или
другой нейрон) с off-центром возбуждается при световой стимуляции
периферии и тормозится при освещении центра своего рецептивного поля
(см. рисунок).[2]
Фоторецепторы, которые включены в состав рецептивных полей нескольких
ганглиозных клеток, способны как возбуждать, так и тормозить
постсинаптические нейроны (англ. postsynaptic neurons), поскольку они высвобождают нейротрансмиттер глутамат
на своих синапсах, что может способствовать как деполяризации, так и
гиперполяризации мембранного потенциала клетки, в зависимости от того,
какие именно ионные каналы открываются нейротрансмиттером. Организация рецептивного поля по принципу центр-периферия позволяет ганглиозным клеткам передавать информацию
не только о том, освещены ли фоторецепторные клетки, но также и о
различиях в параметрах возбуждения подобных клеток, расположенных в
центре и на периферии рецептивного поля. Последнее даёт возможность
ганглиозным клеткам посылать нейронам более высоких синаптических
уровней информацию о контрастности изображения. Размер рецептивного поля
влияет на пространственную частоту (англ. spatial frequency)
визуальной информации: небольшие рецептивные поля активируются
сигналами с высокими пространственными частотами и тонкой детализацией
изображения; большие рецептивные поля — сигналами с низкими
пространственными частотами и плохой детализацией. Рецептивные поля
ганглиозных клеток сетчатки передают информацию о дискретности
распределения света, падающего на сетчатку, а это зачастую позволяет
обнаруживать краевые части визуальных объектов. При адаптации к темноте
инактивируется периферийная зона рецептивного поля, но его активная
часть, а следовательно, площадь суммации сигналов и совокупная
чувствительность, могут реально возрасти вследствие ослабления взаимного
горизонтального торможения центра и периферии рецептивного поля.[3]
Как правило, рецептивные поля лучше реагируют на движущиеся объекты —
такие как светлое или тёмное пятно, пересекающее поле от центра к
периферии (или в противоположном направлении), а также на контуры
объектов — вследствие нарушения равномерности в распределении света по
поверхности поля. Диаметр центральной части рецептивного поля
ганглионарной клетки сетчатки совпадает с протяжённостью её дендритов,
тогда как площадь периферии рецептивного поля определяется амакринными
клетками, утанавливающими связь данной ганглионарной клетки со
множеством биполярных клеток. Кроме того, амакринные клетки могут не
допускать передачи сигналов в ганглионарную клетку от периферии её
рецептивного поля, тем самым усиливая доминирование реакции центра
рецептивного поля («включённый центр и выключенная периферия» — англ. “on-center, off-periphery”). Ганглионарная клетка сетчатки кролика возбуждается при движении светового пятна в «предпочитаемом» (англ. «preferred») направлении и не реагирует, если направление является противоположным («нулевым», англ. «null»).[4][5]
Ганглиозные клетки, способные различать направление движения, найдены
также в сетчатке кошки, земляной белки, голубя. Считается, что
обнаруженные свойства рецептивных полей ганглиозных клеток связаны с
особеннностями сложных механизмов торможения, действующих в сетчатке.[6][7][8]
[править] Латеральное коленчатое тело
На более высоких уровнях зрительной системы группы ганглионарных
(ганглиозных) клеток формируют рецептивные поля нейронов подкоркового
зрительного центра — латерального (наружного) коленчатого тела.
Рецептивные поля напоминают таковые ганглионарных клеток, с
антагонистической системой «центр-периферия»; здесь также имеются
нейроны с on- или off- центрами (приблизительно в равном количестве). [9]
[править] Зрительная кора больших полушарий
Рецептивные поля нейронов зрительной зоны коры крупнее по размерам и
имеют большую избирательность по отношению к визуальным стимулам, нежели
ганглиозные клетки сетчатки или нейроны латерального коленчатого тела. Хьюбел и Визель (например, Hubel, 1963) подразделили рецептивные поля корковых нейронов зрительной системы на «простые», «сложные» и «сверхсложные».[9]«Простые» рецептивные поля имеют удлинённую форму, к примеру, с центральной эллипсовидной
зоной возбуждения и антагонистической зоной торможения по периферии
эллипса. Либо они могут быть почти прямоугольными; при этом одна из
длинных сторон прямоугольника является зоной возбуждения, а другая —
антагонистической зоной торможения. Изображения, активирующие нейроны
этих рецептивных полей, должны быть ориентированы определённым образом.
Чтобы возбудить нейрон со «сложным» рецептивных полем, световому
стимулу в виде полоски недостаточно быть правильно ориентированным —
нужно ещё и двигаться, причём в строго определённом направлении. Для
активации корковых нейронов со «сверхсложными» рецептивными
полями зрительному стимулу в виде полоски необходимо обладать всеми
вышеперечисленными свойствами, и к тому же длина этой полоски должна
быть строго определённой.
[править] Экстрастриарная зрительная кора
Экстрастриарная зрительная кора (поля Бродмана 18 и 19) находится за пределами первичной зрительной коры.[2]
Здесь нейроны могут иметь очень большие рецептивные поля, и для их
активации могут потребоваться очень непростые изображения. Например,
рецептивные поля нейронов нижневисочной извилины (англ. inferotemporal cortex),
пересекают среднюю линию зрительного пространства, и эти нейроны
активируются такими сложными визуальными образами, как радиальная
решётка или кисти рук. Также было обнаружено, что нервные клетки
вентральной поверхности веретеновидной извилины (на границе между
затылочной и височной долями), где находится так называемая «зона
распознавания лиц» (англ. fusiform face area), реагируют, в основном, на изображения лиц.[10] Это важное открытие было получено с помощью технологии функциональной магнитно-резонансной томографии. Позднее оно было подтверждено на уровне исследования нервных клеток.[11]
Подобным способом проводятся поиски других специфических зон зрительной
коры; например, имеются относительно недавние публикации, полагающие,
что так называемая парагиппокампальная навигационная зона (англ. parahippocampal place area)
может быть отчасти специализрована к распознаванию зданий. Кстати, в
одном из последних исследований высказывается предположение, что «зона
распознавания лиц» веретеновидной извилины, возможно, не только
выполняет функцию, отражённую в её наименовании, но и вообще служит для
различения отдельных частей целого.
Источник

