
Амакриновые клетки сетчатки и латеральное торможение
Амакриновые клетки. Ганглиозные клетки сетчатки
В настоящее время морфологическими или гистохимическими методами идентифицированы около 30 типов амакриновых клеток. Функции некоторых из них охарактеризованы, и все они отличаются друг от друга. Один тип амакриновых клеток является частью прямого пути палочкового зрения, т.е. от палочки к биполярным клеткам, затем к амакриновым и, наконец, к ганглиозным клеткам.
Амакриновые клетки другого типа активно реагируют в начале непрерывного зрительного сигнала, но ответ быстро исчезает. Некоторые амакриновые клетки, наоборот, мощно реагируют на выключение зрительного сигнала, но их ответ также быстро прекращается.
Амакриновые клетки еще одного типа реагируют и на включение, и на выключение света, просто сигнализируя об изменении освещения, независимо от его направления. Есть амакриновые клетки, реагирующие на движение пятна света по сетчатке в определенном направлении; о таких клетках говорят, что они чувствительны к направлению.
В известном смысле большинство амакриновых клеток являются вставочными нейронами, которые помогают анализировать зрительные сигналы, прежде чем они покинут сетчатку.
Ганглиозные клетки
Каждая сетчатка содержит около 100 млн палочек и 3 млн колбочек; однако количество ганглиозных клеток — лишь около 1,6 млн. Таким образом, в среднем 60 палочек и 2 колбочки конвергируют на каждую ганглиозную клетку и волокно зрительного нерва, идущее от ганглиозной клетки к мозгу.
Однако существуют большие различия между периферической и центральной областями сетчатки. По мере приближения к ямке все меньше палочек и колбочек конвергируют на каждое зрительное волокно; кроме того, и палочки, и колбочки становятся тоньше. Эти эффекты постепенно увеличивают остроту зрения в центральной сетчатке. В самом центре — в области центральной ямки — есть только тонкие колбочки (примерно 35000) и совсем нет палочек. К тому же, как показано справа на рисунке, количество волокон зрительного нерва, выходящих из этой части сетчатки, почти равно числу колбочек. Это объясняет высокую степень остроты зрения в центральной сетчатке по сравнению с гораздо меньшей остротой на периферии.
Другим различием между периферией и центром сетчатки является гораздо более высокая чувствительность периферической сетчатки к слабому свету. Отчасти это объясняется тем, что чувствительность палочек в 30-300 раз выше, чем колбочек. Однако эффект значительно усиливается в связи с конвергенцией примерно 200 палочек на одну ганглиозную клетку в периферических областях сетчатки, поэтому сигналы от палочек суммируются, дополнительно усиливая интенсивность стимуляции периферических ганглиозных клеток и исходящих от них волокон зрительных нервов.
Различают три группы ганглиозных клеток, которые обозначают как W-, Х- и Y-клетки. Каждая группа выполняет свою функцию.
Передача палочкового зрения W-клетками. W-клетки составляют примерно 40% общего числа ганглиозных клеток. Они маленькие (диаметром около 10 мкм) и передают сигналы по соответствующим им волокнам зрительного нерва с низкой скоростью, равной около 8 м/сек. Эти ганглиозные клетки возбуждаются в основном от палочек, передающих к ним сигналы через малые биполярные и амакриновые клетки. W-клетки имеют широкие рецептивные поля в сетчатке, т.к. их дендриты широко распространяются во внутреннем слое сетчатки, получая сигналы от обширных областей.
На основании гистологических и физиологических экспериментов показано, что W-клетки, по-видимому, особенно чувствительны к восприятию направленного движения в поле зрения и очень важны для нашего грубого палочкового зрения в условиях темноты.
Передача зрительного образа и цвета Х-клетками. Подавляющее большинство ганглиозных клеток (55%) являются Х-клетками. Они имеют средний диаметр (в пределах от 10 до 15 мкм) и передают сигналы по своим волокнам зрительного нерва со скоростью около 14 м/сек.
Рецептивные поля Х-клеток небольшие, поскольку их дендриты не имеют широкого распространения в сетчатке. В связи с этим сигналы Х-клеток отражают дискретные участки сетчатки. Следовательно, в основном через Х-клетки передаются тонкие детали зрительного образа. Кроме того, поскольку каждая Х-клетка получает сигналы, по крайней мере, от одной колбочки, эти клетки, вероятно, отвечают за все цветовое зрение.
Функция Y-клеток — передача информации о мгновенных изменениях в зрительном образе. Y-клетки — самые большие из всех ганглиозных клеток (диаметром до 35 мкм). Они проводят сигналы к мозгу со скоростью 50 м/сек и выше. Среди ганглиозных клеток они самые малочисленные (около 5% общего количества) и имеют разветвленные дендриты, следовательно, собирают сигналы от обширных областей сетчатки.
Как многие из амакриновых клеток, Y-клетки реагируют на быстрые изменения в зрительном образе (быстрые движения или быстрые изменения освещения), посылая импульсные разряды, длительность которых составляет лишь доли секунды. Эти ганглиозные клетки, вероятно, почти немедленно информируют центральную нервную систему о появлении любого нового зрительного явления в поле зрения, но без высокой точности его локализации, обеспечивая лишь соответствующие сигналы, заставляющие глаза двигаться по направлению к возбуждающему объекту.
— Также рекомендуем «Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке»
Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»:
1. Каскад усиления в сетчатке. Фотохимия цветового зрения
2. Световая и темновая адаптация. Механизмы световой и темновой адаптации
3. Острота зрения. Определение расстояния до объекта глазами
4. Слепота на отдельные цвета. Функция нейронов сетчатки
5. Зрительный путь от колбочек. Нейромедиаторы нейронов сетчатки
6. Функция горизонтальных клеток сетчатки. Возбуждение и торможение биполярных клеток
7. Амакриновые клетки. Ганглиозные клетки сетчатки
8. Возбуждение ганглиозных клеток. Роль латерального торможения в сетчатке
9. Зрительные пути. Дорсолатеральное коленчатое ядро таламуса
10. Зрительная кора. Строение первичной зрительной коры
Источник
Сетчатка (лат. retína) — внутренняя оболочка глаза, являющаяся периферическим отделом зрительного анализатора; содержит фоторецепторные клетки, обеспечивающие восприятие и преобразование электромагнитного излучения видимой части спектра в нервные импульсы, а также обеспечивает их первичную обработку.
В горизонтальных, биполярных и амакриновых клетках переработка сигнала происходит путем медленных изменений мембранного потенциала в синапсах под влиянием непрерывного выделения медиатора из фоторецептора; потенциалы действия здесь не генерируются. При взаимодействии фоторецептора с квантом света и соответствующей гиперполяризации снижается интенсивность образования медиатора и как следствие деполяризация биполярной клетки. Это приводит к тому, что на противоположной стороне биполярной клетки выделяется второй медиатор, вызывающий деполяризацию мембраны ганглиозной клетки и генерацию потенциалов действия.
Синаптические окончания фоторецепторов конвергируют на биполярные нейроны сетчатки (первый нейрон). При этом фоторецепторы центральной ямки связаны с одним биполяром. Аксоны биполярных клеток в свою очередь конвергируют на ганглиозные клетки (второй нейрон).
В результате на каждую ганглиозную клетку могут конвергировать около 140 палочек и 6 колбочек, при этом, чем ближе к желтому пятну, тем меньше фоторецепторов конвергирует на одну клетку. В области желтого пятна конвергенция почти не осуществляется и количество колбочек почти равно количеству биполярных и ганглиозных клеток.
Периферия сетчатки отличается большой чувствительностью к слабому свету. Это обусловлено тем, что до 600 палочек конвергируют здесь через биполярные клетки на одну и ту же ганглиозную клетку.
В одних ганглиозных клетках учащение фоновых разрядов происходит на включение света (on-ответ), в других – на выключение света (off-ответ). Реакция ганглиозной клетки может быть обусловлена и спектральным составом света.
В сетчатке кроме вертикальных существуют также латеральные связи. Латеральное взаимодействие рецепторов осуществляется горизонтальными клетками.
Биполярные и ганглиозные клетки взаимодействуют между собой за счет многочисленных латеральных связей, образованных коллатералями дендритов и аксонов самих клеток, а также с помощью амакриновых клеток.
Горизонтальные клетки сетчатки обеспечивают регуляцию передачи импульсов между фоторецепторами и биполярами, регуляцию цветовосприятия и адаптации глаза к различной освещенности. В течение всего периода освещения горизонтальные клетки генерируют положительный потенциал – медленную гиперполяризацию, названную S-потенциалом. По характеру восприятия световых раздражений горизонтальные клетки делят на два типа: 1) L-тип, в котором S-потенциал возникает при действии любой волны видимого света; 2) С-тип, или «цветовой» тип, в котором знак отклонения потенциала зависит от длины волны. Так, красный свет может вызвать их деполяризацию, а синий – гиперполяризацию.
Горизонтальные, а также амакриновые клетки называют тормозными нейронами, так как они обеспечивают латеральное торможение между биполярными или ганглиозными клетками.
Аксоны ганглиозных клеток собираются в диске зрительного нерва (слепого пятна), образуя ствол зрительного нерва.
Источник
Функция горизонтальных клеток сетчатки. Возбуждение и торможение биполярных клетокГоризонтальные клетки, показанные на рисунке, обеспечивают связь между синаптическими телами палочек и колбочек и дендритами биполярных клеток. На выходе горизонтальные клетки всегда тормозные. Следовательно, эта латеральная связь обеспечивает феномен латерального торможения, характерный для всех других сенсорных систем и важный для повышения надежности проведения визуального образа с соответствующим зрительным контрастом. Этот феномен демонстрируется на рисунке, где показано небольшое пятно света, сфокусированное на сетчатке. Зрительный путь от самой центральной области, куда попадает световой луч, возбуждается, тогда как рядом лежащая область тормозится. Другими словами, горизонтальные клетки путем латерального торможения в окружающих областях останавливают диффузное распространение сигнала по сетчатке, которое могло бы возникнуть в связи с наличием широкого ветвления дендритов и аксонов в слоях сетчатки. Это важно для четкого выделения контрастных границ в зрительном образе. Возможно, что некоторые амакриновые клетки также обеспечивают дополнительное латеральное торможение и дальнейшее усиление зрительного контраста во внутреннем слое сетчатки. Возбуждение и торможение биполярных клетокПротивоположные сигналы (возбуждающие и тормозные) в зрительном пути обеспечиваются двумя типами клеток: (1) деполяризующимися биполярными клетками (2) гиперполяризующимися биполярными клетками. Это значит, что при возбуждении палочек и колбочек некоторые биполярные клетки деполяризуются, тогда как другие гиперполяризуются. Существуют два возможных объяснения этого различия. Согласно одному из них имеются биполярные клетки двух типов, которые по-разному реагируют на выделяемый палочками и колбочками медиатор глутамат: одни деполяризуются, другие гиперполяризуются. Другим возможным объяснением является то, что одни биполярные клетки получают сигнал непосредственно от палочек и колбочек, тогда как другие получают этот же сигнал через горизонтальные клетки. Поскольку горизонтальные клетки тормозные, их действие приводит к противоположной реакции биполярных клеток. Независимо от механизма разнонаправленных реакций, важность этого феномена состоит в том, что одна половина биполярных клеток проводит положительные сигналы, а другая — отрицательные. Как мы увидим далее, оба типа сигналов — и положительный, и отрицательный — используются при проведении зрительной информации в мозг. Другим важным аспектом этого реципрокного отношения между деполяризующимися и гиперполяризующимися биполярными клетками является обеспечение второго механизма латерального торможения дополнительно к механизму, связанному с горизонтальными клетками. Поскольку разные типы биполярных клеток лежат рядом друг с другом, это способствует появлению разделяющих контрастных границ в зрительном образе, даже если граница лежит точно между двумя прилежащими фоторецепторами. В противоположность этому механизм латерального торможения, обеспечиваемый горизонтальными клетками, действует на гораздо большем расстоянии.
— Также рекомендуем «Амакриновые клетки. Ганглиозные клетки сетчатки» Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»: |
Источник
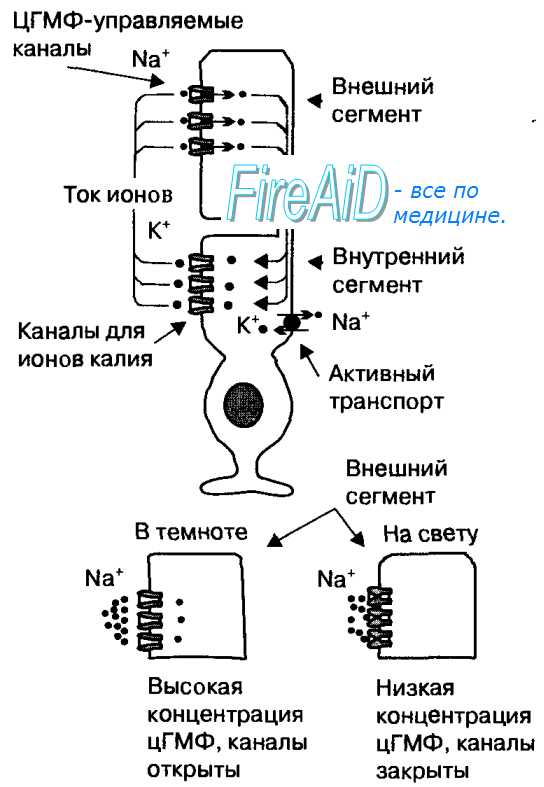
Возбуждение ганглиозных клеток. Роль латерального торможения в сетчаткеСпонтанные постоянные потенциалы действия в ганглиозных клетках. Именно от ганглиозных клеток длинные волокна зрительного нерва направляются в мозг. В связи с большим расстоянием электротонический способ проведения, используемый палочками, колбочками и биполярными клетками в пределах сетчатки, больше не годится; вместо этого ганглиозные клетки проводят свои сигналы в виде периодических потенциалов действия. Более того, даже при отсутствии стимуляции они посылают постоянные импульсы с частотой 5-40 в секунду. Зрительные сигналы, в свою очередь, накладываются на эту фоновую активность ганглиозных клеток. Передача информации об изменении освещенности. Ответы «on—off» и «off—on». Как уже указывалось, многие ганглиозные клетки возбуждаются специфически при изменениях освещенности. Это демонстрируется изменениями импульсной активности на рисунке. На верхней кривой представлена импульсная активность, которая сразу при включении света на долю секунды резко усиливается, но в следующую долю секунды снижается. Нижняя запись отражает активность ганглиозной клетки, расположенной латерально по отношению к пятну света; эта клетка явно тормозится при включении света из-за латерального торможения. При выключении света происходят противоположные явления. Таким образом, эти записи отражают так называемые ответы «on-off» и «off-on». Противоположные направления этих ответов связаны, соответственно, с деполяризующимися и гиперполяризующимися биполярными клетками, а нестационарная природа ответов, по крайней мере частично, генерируется амакриновыми клетками, многие из которых имеют такие же нестационарные реакции. Способность глаз определять изменение освещенности весьма развита и в периферической, и в центральной сетчатке. Например, очень маленький комар, летящий через поле зрения, обнаруживается мгновенно. И наоборот, тот же комар, если он тихо сидит, остается ниже порога зрительного восприятия. Роль латерального торможенияМногие ганглиозные клетки реагируют главным образом на контрастные границы в поле зрения. Поскольку, видимо, это главный способ передачи в мозг информации об особенностях рассматриваемого объекта, попробуем объяснить, как это происходит. Если на сетчатку падает ровный свет, т.е. все фоторецепторы равномерно стимулируются падающим светом, ганглиозные клетки контрастного типа не стимулируются и не тормозятся. Это связано с тем, что сигналы, проводимые непосредственно от фоторецепторов через деполяризующиеся биполярные клетки, являются возбуждающими, тогда как сигналы, проводимые латералъно через гиперполяризующиеся биполярные клетки, как и через горизонтальные клетки, являются главным образом тормозными. Таким образом, прямой возбуждающий сигнал, идущий по одному пути, вероятно, должен нейтрализоваться тормозными сигналами, проходящими через латеральный путь. Один из таких контуров показан на рисунке, в верхней части которого — три фоторецептора. Центральный рецептор возбуждает деполяризующуюся биполярную клетку. Два боковых рецептора связаны с той же биполярной клеткой через тормозные горизонтальные клетки, которые нейтрализуют прямой возбуждающий сигнал, если три рецептора возбуждаются светом одновременно.
Теперь рассмотрим, что происходит при наличии контрастной границы в видимом образе. Обратившись вновь к рисунку, предположим, что центральный фоторецептор стимулируется ярким пятном света, тогда как один из двух латеральных рецепторов находится в темноте. Яркое пятно света возбуждает прямой путь через биполярную клетку. Пребывание одного из латеральных рецепторов в темноте ведет к тому, что состояние одной из горизонтальных клеток не изменяется, т.е. она не стимулируется. Следовательно, эта клетка не тормозит биполярную клетку. Таким образом, там, где есть зрительные контрасты, сигналы прямого и латерального путей усиливают друг друга. В общем, функция механизма латерального торможения в глазу — та же, что и в большинстве других сенсорных систем: обеспечить выявление и усиление контраста. Одиночная ганглиозная клетка может стимулироваться либо многими колбочками, либо немногими. Когда все три типа колбочек (красные, синие и зеленые) стимулируют одну и ту же ган-глиозную клетку, сигнал, передаваемый через нее, — один и тот же для любого цвета спектра. Следовательно, сигнал от данной ганглиозной клетки не играет никакой роли в определении цвета, т.е. сигнал «белый». С другой стороны, некоторые из ганглиозных клеток возбуждаются колбочкой одного цветового типа и тормозятся колбочкой другого цветового типа. Это часто бывает в отношении красных и зеленых колбочек, когда красные вызывают возбуждение, а зеленые — торможение или наоборот. Такой же тип реципрокных взаимовлияний существует между синими колбочками, с одной стороны, и сочетанием красных и зеленых колбочек (оба типа возбуждаются желтым цветом), с другой стороны. Это ведет к реципрокным отношениям (возбуждение — торможение) между синим и желтым цветами. Механизм такого противоположного влияния цветов следующий. Колбочки одного цветового типа возбуждают ганглиозную клетку по прямому возбуждающему пути через деполяризующуюся биполярную клетку, тогда как колбочки другого цветового типа тормозят эту ганглиозную клетку непрямым тормозным путем через гиперполяризующуюся биполярную клетку. Важность этих цветоконтрастных механизмов состоит в том, что они отражают способ различения цветов на уровне самой сетчатки. Таким образом, каждый цветоконтрастный тип ганглиозной клетки возбуждается одним цветом, но тормозится «оппонентным» цветом. Следовательно, анализ цвета начинается в сетчатке и не является полностью функцией головного мозга. — Также рекомендуем «Зрительные пути. Дорсолатеральное коленчатое ядро таламуса» Оглавление темы «Физиология сетчатки. Проводящие зрительные пути»: |
Источник

